
Основы генетики.
Гибридологический метод генетического анализа, его основные принципы.
Генетический анализ — исследование генотипа отдельных особей, групп особей и генетической структуры популяций, в том числе линий, штаммов, сортов, пород и т.д.
Гибридологический анализ, способ изучения наследственных свойств организма путём скрещивания (гибридизации) его с родственной формой и последующим анализом признаков потомства. Гибридологический анализ впервые применил Г. Мендель (1865) для изучения механизма передачи наследственных задатков (генов) от родителей потомкам и для изучения взаимодействия генов у одного и того же организма. В основе гибридологического анализа лежит способность к рекомбинации, т. е. перераспределению генов при образовании гамет, что приводит к возникновению новых сочетаний генов. По этим сочетаниям, которые проявляются в потомстве гибридной особи с определённой частотой, можно судить о генотипе родительской формы, а по генотипу родительской формы можно предсказывать генотип потомства. Так, генотип особи, гибридной по паре аллелей, одна из которых — доминантная А, другая — рецессивная а, можно представить как Аа. Внешне, т. е. фенотипически, такая форма (гетерозигота) не отличается от формы с генотипом АА (гомозигота). Гибрид (Аа) формирует гаметы двух типов, каждый из которых несёт аллель А или аллель а. Таким образом, гаметы никогда не бывают гибридными. С помощью различных видов скрещивания можно выявить, сколько типов гамет по данному гену формирует организм, и определить его генотип. Если у анализируемой формы (Аа) возможно самооплодотворение (что часто встречается у растений), схематично это будет выглядеть так: ♂ (А+а) ´ ♀ (А+а) (АА + Аа + Аа + аа. При этом в потомстве с определённой частотой появляется новая форма — аа.
Если самооплодотворения нет, генотип исходной формы выявляют, скрещивая в разных комбинациях её потомков ( «брат ´ сестра») и анализируя «внучатое» поколение. Другой способ выявления гибридного состояния — анализирующее скрещивание: скрещивание предполагаемого гибрида с рецессивной родительской формой. Гибридологический анализ играет важную роль в селекционной практике и племенном деле, т.к. позволяет судить о тождестве фенотипа и генотипа. Здесь гибридологический анализ находит применение в форме «анализа производителей по потомству» с целью выявления у производителей скрытых нежелательных генов. Гибридологический анализ применяется также при составлении хромосомных карт. Знание генного состава хромосомы позволяет путём специальных скрещиваний вводить в геном определённую хромосому или группу генов и создавать формы с нужным генотипом. Этот метод широко применяется в растениеводстве. Гибридологическим анализом пользуются при изучении взаимодействия генов в первом гибридном поколении (тесты на комплементацию). Гибридологический анализ является главным методом генетического анализа.
Гибридологический анализ, основан на следующих принципах.
1. Использование в качестве исходных особей (родителей), форм, не дающих расщепления при скрещивании, т.е. константных форм.
2. Анализ наследования отдельных пар альтернативных признаков, то есть признаков, представленных двумя взаимоисключающими вариантами.
3. Количественный учет форм, выщепляющихся в ходе последовательных скрещиваний и использование математических методов при обработке результатов.
4. Индивидуальный анализ потомства от каждой родительской особи.
5. На основании результатов скрещивания составляется и анализируется схема скрещиваний.
Понятие о гене, строение, функции и свойства гена.
Ген — структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения.
Термин «ген» был введён в употребление в 1909 году датским ботаником Вильгельмом Йогансеном. Изучением генов занимается наука генетика, родоначальником которой считается Грегор Мендель, который в 1865 году опубликовал результаты своих исследований о передачи по наследству признаков при скрещивании гороха. Сформулированные им закономерности впоследствии назвали Законами Менделя.
Среди ученых нет единого мнения под каким углом рассматривать ген. Одни ученые его рассматривают как информационную наследственную единицу, а единицей естественного отбора является вид, группа, популяция или отдельный индивид. Другие ученные, как например Ричард Докинз в своей книге «Эгоистичный ген», рассматривает ген как единицу естественного отбора, а сам организм — как машину для выживания генов.
В настоящее время, в молекулярной биологии установлено, что гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК. Эти и другие функциональные молекулы определяют развитие, рост и функционирование организма.
В то же время, каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК, которые принимают непосредственное участие в регулировании проявления гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания, кодирующей белок, или начала последовательности РНК, так и на расстоянии многих миллионов пар оснований (нуклеотидов). Таким образом, понятие гена не ограничено только кодирующим участком ДНК, а представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.
Изначально термин ген появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Большинство исследователей считали, что такими носителями могут быть только белки, так как их строение (20 аминокислот) позволяет создать больше вариантов, чем строение ДНК, которое составлено всего из четырёх видов нуклеотидов. Позже было экспериментально доказано, что именно ДНК включает в себя наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии.
Гены могут подвергаться мутациям — случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Однако, далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными.
Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие в себя азотистые основания: аденин (А) или тимин (Т) или цитозин (Ц) или гуанин (Г), пятиатомный сахар-пентозу-дезоксирибозу, по имени которой и получила название сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов.
В процессе реализации наследственной информации, заключенной в гене, проявляется целый ряд его свойств. Определяя возможность развития отдельного качества, присущего данной клетке или организму, ген характеризуется дискретностью действия (от лат. discretus — разделенный, прерывистый), прерывностью (интроны и экзоны). Дискретность наследственного материала, предположение о которой высказал еще Г. Мендель, подразумевает делимость его на части, являющиеся элементарными единицами, — гены. В настоящее время ген рассматривают как единицу генетической функции. Он представляет собой минимальное количество наследственного материала, которое необходимо для синтеза тРНК, рРНК или полипептида с определенными свойствами. Ген несет ответственность за формирование и передачу по наследству отдельного признака или свойства клеток, организмов данного вида. Кроме того, изменение структуры гена, возникающее в разных его участках, в конечном итоге приводит к изменению соответствующего элементарного признака.
Ввиду того что в гене заключается информация об аминокислотной последовательности определенного полипептида, его действие является специфичным. Однако в некоторых случаях одна и та же нуклеотидная последовательность может детерминировать синтез не одного, а нескольких полипептидов. Это наблюдается в случае альтернативного сплайсинга у эукариот и при перекрывании генов у фагов и прокариот. Очевидно, такую способность следует оценить как множественное, или плейотропное, действие гена (хотя традиционно под плейотропным действием гена принято понимать участие его продукта — полипептида — в разных биохимических процессах, имеющих отношение к формированию различных сложных признаков). Например, участие фермента в ускорении определенной реакции, которая является звеном нескольких биохимических процессов, делает зависимыми результаты этих процессов от нормального функционирования гена, кодирующего этот белок. Нарушение реакции A>B, катализируемой белком б, в результате мутации гена ведет к выключению последующих этапов формирования признаков D и E.
б D
A B
C
E
Определяя возможность транскрибирования мРНК для синтеза конкретной полипептидной цепи, ген характеризуется дозированностью действия, т.е. количественной зависимостью результата его экспрессии от дозы соответствующего аллеля этого гена.
Свойства гена:
- дискретность — несмешиваемость генов;
- стабильность — способность сохранять структуру;
- лабильность — способность многократно мутировать;
- множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
- аллельность — в генотипе диплоидных организмов только две формы гена;
- специфичность — каждый ген кодирует свой признак;
- плейотропия — множественный эффект гена;
- экспрессивность — степень выраженности гена в признаке;
- пенетрантность — частота проявления гена в фенотипе;
- амплификация — увеличение количества копий гена.
Моногибридное, дигибридное, полигибридные и другие типы скрещиваний.
Моногибридным называется скрещивание, при котором родительские формы отличаются друг от друга по одной паре контрастных, альтернативных признаков.
Признак — любая особенность организма, т. е. любое отдельное его качество или свойство, по которому можно различить две особи. Совокупность всех признаков организма, начиная с внешних и кончая особенностями строения и функционирования клеток, тканей и органов, называется фенотипом. Этот термин может употребляться и по отношению к одному из альтернативных признаков.
Признаки и свойства организма проявляются под контролем наследственных факторов, т. е. генов. Совокупность всех генов организма называют генотипом.
Примерами моногибридного скрещивания, проведенного Г. Менделем, могут служить скрещивания гороха с такими хорошо заметными альтернативными признаками, как пурпурные и белые цветки, желтая и зеленая окраска незрелых плодов (бобов), гладкая и морщинистая поверхность семян, желтая и зеленая их окраска и др.
Единообразие гибридов первого поколения (первый закон Менделя). При скрещивании гороха с пурпурными и белыми цветками Мендель обнаружил, что у всех гибридных растений первого поколения (F1) цветки оказались пурпурными. При этом белая окраска цветка не проявлялась (рис. 1).
Мендель установил также, что все гибриды F1 оказались единообразными (однородными) по каждому из семи исследуемых им признаков.
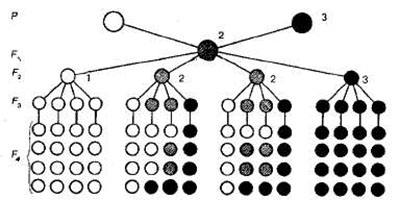
Рис.1. Схема моногибридного скрещивания: I — гомозиготные особи с доминантным признаком; 2 — гетерозиготные особи с доминантным или промежуточным признаком; 3 — гомозиготные особи с рецессивным признаком.
Следовательно, у гибридов первого поколения из пары родительских альтернативных признаков проявляется только один, а признак другого родителя как бы исчезает. Явление преобладания у гибридов F1 признаков одного из родителей Мендель назвал доминированием, а соответствующий признак — доминантным. Признаки, не проявляющиеся у гибридов F1 он назвал рецессивными.
Поскольку все гибриды первого поколения единообразны, это явление было названо К. Корренсом первым законам Менделя, или законом единообразия гибридов первого поколения, а также правилом доминирования.
Закон расщепления (второй закон Менделя). Из гибридных семян гороха Мендель вырастил растения, которые подверг самоопылению, и образовавшиеся семена вновь высеял. В результате было получено второе поколение гибридов, или гибриды F2. Среди последних обнаружилось расщепление по каждой паре альтернативных признаков в соотношении примерно 3:1, т. е. три четверти растений имели доминантные признаки (пурпурные цветки, желтые семена, гладкие семена и т. д.) и одна четверть — рецессивные (белые цветки, зеленые семена, морщинистые семена и т. д.). Следовательно, рецессивный признаку гибрида F1 не исчез, а только был подавлен и вновь проявился во втором поколении. Это обобщение позднее было названо вторым законом Менделя, или законом расщепления.
Гомозиготные и гетерозиготные особи. Чтобы выяснить, как будет осуществляться наследование признаков в третьем, четвертом и последующих поколениях, Мендель путем самоопыления вырастил гибриды этих поколений и проанализировал полученное потомство. Он выяснил, что растения, обладающие рецессивными признаками (например, белые цветки), в следующих поколениях (F3 F4 и т. д.), воспроизводят потомство только с белыми цветками (см. рис. 1).
Иначе вели себя гибриды второго поколения, обладающие доминантными признаками (например, пурпурными цветками). Среди них при анализе потомства Мендель обнаружил две группы растений, внешне совершенно неразличимых по каждому конкретному признаку.
Первая группа, составляющая 1/3 от общего числа растений с доминантным признаком, далее не расщеплялась, т. е. во всех последующих поколениях у них обнаруживалась только пурпурная окраска цветков. Оставшиеся ⅔ растений второго поколения в F3, снова давали расщепление такое же, как в F2 т. е. на три растения с пурпурными цветками появлялось одно с белыми.
Особи, которые не дают в потомстве расщепления и сохраняют свои признаки в «чистом» виде, называют гомозиготными, а те, у которых в потомстве происходит расщепление, —гетерозиготными.
Таким образом, Менделем впервые было установлено, что растения, сходные по внешним признакам, могут обладать различными наследственными свойствами.
Аллелизм. Для установления причины расщепления, причем в строго определенных численных отношениях доминантных и рецессивных признаков, следует вспомнить, что связь между поколениями при половом размножении осуществляется через половые клетки (гаметы). Очевидно, гаметы несут материальные наследственные задатки, или факторы, определяющие развитие того или иного признака. Эти факторы позже и были названы генами.
В соматических клетках диплоидного организма эти задатки являются парными: один получен от отцовского организма, а другой — от материнского. Мендель предложил обозначать доминантные наследственные задатки заглавной буквой (например,А), а соответствующие им рецессивные задатки прописной буквой (а). Пару генов, определяющих альтернативные признаки, называют аллеломорфной парой, а само явление парности — алле-лизмом.
Каждый ген имеет два состояния — А и а, поэтому они составляют одну пару, а каждого из членов пары называют аллелем. Таким образом, гены, расположенные в одних и тех же локусах (участках) гомологических хромосом и определяющие альтернативное развитие одного и того же признака, называются аллельными. Например, пурпурная и белая окраска цветка гороха является доминантным и рецессивным признаками соответственно двум аллелям (А и а) одного гена. Благодаря наличию двух аллелей возможны два состояния организма: гомо- и гетерозиготные. Если организм содержит одинаковые аллели конкретного гена (АА или аа), то он называется гомозиготным по данному гену (или признаку), а если разные (Аа) — то гетерозиготным. Следовательно, аллель — это форма существования гена.
Примером трехаллельного гена является ген, определяющий у человека систему группы крови АВ0. Аллелей бывает и больше: для гена, контролирующего синтез гемоглобина человека, их известно много десятков.
Статистический анализ расщепления. Представим результаты опытов Менделя по моногибридному скрещиванию гороха в виде схемы (рис. 2). Символы Р, F1, F2 и т- д. обозначают родительское, 1-е и 2-е поколение соответственно, знак умножения указывает скрещивание, символ о* обозначает мужской пол, a Q — женский. Из схемы видно, что в родительском поколении (Р) материнская и отцовская формы гомозиготны по исследуемому признаку, поэтому производят гаметы только с аллелем А или только с а.
При оплодотворении эти гаметы образуют зиготу, которая имеет оба аллеля Аа — доминантный и рецессивный. В результате все гибриды F1 единообразны по конкретному признаку, поскольку доминантный аллель А подавляет действие рецессивного аллеля а. Во время образования гамет аллели А и а попадают в них по одному. Следовательно, гибридные организмы способны производить гаметы двух типов, несущие аллели А и а, т. е. являются гетерозиготными.
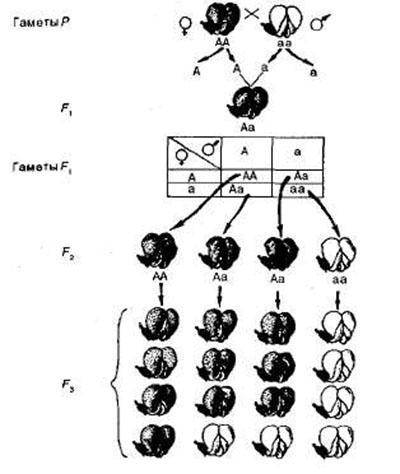
Рис. 2. Наследование пурпурной и белой окраски цветков гороха.
Для облегчения расчета сочетаний разных типов гамет английский генетик Р. Пеннет предложил производить запись в виде решетки, которая и вошла в литературу под назван и ем решетка Пеннета (см. рис. 2). Слева по вертикали располагаются женские гаметы, сверху по горизонтали — мужские. В квадраты решетки вписывают образующиеся сочетания гамет, которые соответствуют генотипам зигот.
При самоопылении в F2 получается расщепление по генотипу в отношении 1АА:2Аа:1аа, т. е. одна четвертая часть гибридов гомозиготны по доминантным аллелям, половина — гетерозиготны и одна четвертая часть — гомозиготны по рецессивным аллелям. Так как генотипам АА и Аа соответствует один и тот же фенотип — пурпурная окраска цветка, расщепление по фенотипу будет следующим; 3 пурпурных: 1 белый. Следовательно, расщепление по фенотипу не совпадает с расщеплением по генотипу.
Теперь легко объяснить, почему гомозиготные белоцветковые растения второго поколения с рецессивными аллелями аа при самоопылении b F3 дают только себе подобных. Такие растения производят гаметы одного типа, и, как следствие, расщепления не наблюдается. Ясно также, что среди пурпурноцветковых 1 /3 доминантных гомозигот (АА) также не будет давать расщепления, а ⅔ гетерозиготных растений (Аа) будут давать b F3 расщепление 3:1, как и у гибридов F2
На основании, анализа результатов моногибридното скрещивания были сформулированы не только первый и второй законы Менделя и правило доминирования, но и правило чистоты гамет.
Правило чистоты гамет. При моногибридном скрещивании в случае полного доминирования у гетерозиготных гибридов (Аа) первого поколения проявляется только доминантный аллель (А); рецессивный же (а) не теряется и не смешивается с доминантным. В F2 как рецессивный, так и доминантный аллели могут проявляться в своем «чистом» виде. При этом аллели не только не смешиваются, но и не претерпевают изменений после совместного пребывания в гибридном организме. В результате гаметы, образуемые такой гетерозиготой, являются «чистыми» в том смысле, что гамета А «чиста» и не содержит ничего от аллеля а, а гамета а «чиста» от А. Это явление несмешивания аллелей пары альтернативных признаков в гаметах гибрида получило название правило чистоты гамет. Данное правило, сформулированное У. Бэтсоном, указывает на дискретность гена, несмешиваемость аллелей друг с другом и другими генами. Цитологическая основа правила чистоты гамет и закона расщепления заключается в том, что гомологичные хромосомы и локализованные в них гены, контролирующие альтернативные признаки, распределяются по разным гаметам.
Анализирующее скрещивание. При полном доминировании судить о генотипе организма по его фенотипу невозможно, поскольку и доминантная гомозигота (АА), и гетерозигота (Аа) обладают фено-типически доминантным признаком. Для того чтобы отличить доминантную гомозиготу от гетерозиготной, используют метод, называемый анализирующим скрещиванием, т. е. скрещивание исследуемого организма с организмом, гомозиготным по рецессивным аллелям. В этом случае рецессивная форма (аа) образует только один тип гамет с аллелем а, что позволяет проявиться любому из двух аллелей исследуемого признака уже в первом поколении.
Например, у плодовой мухи дрозофилы длинные крылья доминируют над зачаточными. Особь с длинными крыльями может быть гомозиготной (LL) или гетерозиготной (Ll). Для установления ее генотипа надо провести анализирующее скрещивание между этой мухой и мухой, гомозиготной по рецессивным аллелям. Если у всех потомков от этого скрещивания будут длинные крылья, то особь с неизвестным генотипом гомозиготна по доминантным аллелям (LL), Если же в первом поколении произойдет расщепление на доминантные и рецессивные формы в отношении 1:1, то можно сделать вывод, что исследуемый организм является гетерозиготным.
Таким образом, по характеру расщепления можно проанализировать генотип гибрида, типы гамет, которые он образует, и их соотношение. Поэтому анализирующее скрещивание является очень важным приемом генетического анализа и широко используется в генетике и селекции.
Скрещивание, при котором родительские формы отличаются по двум парам альтернативных признаков (по двум парам аллелей), называется дигибридным. Гибриды, гетерозиготные по двум генам, называют дигетерозиготными, а в случае отличия их по трем и многим генам —три- и полигетерозиготными соответственно.
Результаты дигибридного и полигибридного скрещивания зависят от того, располагаются гены, определяющие рассмотренные признаки, в одной хромосоме или в разных.
Независимое наследование (третий закон Менделя). Для дигибридного скрещивания Мендель использовал гомозиготные растения гороха, различающиеся одновременно по двум парам признаков. Одно из скрещиваемых растений имело желтые гладкие семена, другое — зеленые морщинистые (рис. 3).
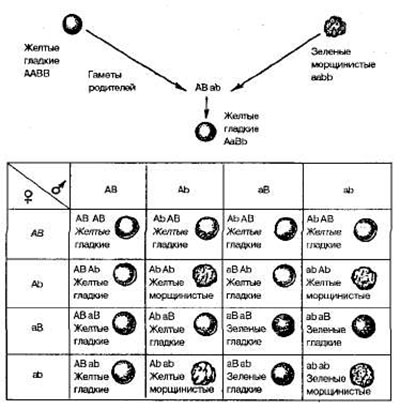
Рис 3. Дигибридное скрещивание растений гороха, различающихся по форме и окраске семян
Все гибриды первого поколения этого скрещивания имели желтые гладкие семена. Следовательно, доминирующими оказались желтая окраска семян над зеленой и гладкая форма над морщинистой. Обозначим аллели желтой окраски А, зеленой — а, гладкой формы— В, морщинистой— b. Гены, определяющие развитие разных пар признаков, называются неаллельпыми и обозначаются разными буквами латинского алфавита. Родительские растения в этом случае имеют генотипы АА ВВ и aabb, а генотип гибридов F1 —АаВb, т. е. является дигетерозиготным.
Во втором поколении после самоопыления гибридов F1 в соответствии с законом расщепления вновь появились морщинистые и зеленые семена. При этом наблюдались следующие сочетания признаков: 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых семян. Это соотношение очень близко к соотношению 9:3:3:1.
Чтобы выяснить, как ведет себя каждая пара аллелей в потомстве дигетерозиготы, целесообразно провести раздельный учет каждой пары признаков — по форме и окраске семян. Из 556 семян Менделем получено 423 гладких и 133 морщинистых, а также 416 желтых и 140 зеленых. Таким образом, и в этом случае соотношение доминантных и рецессивных форм по каждой паре признаков свидетельствует о моногибридном расщеплении по фенотипу 3:1. Отсюда следует, что дигибридное расщепление представляет собой два независимо идущих моногибридных расщепления, которые как бы накладываются друг на друга.
Проведенные наблюдения свидетельствуют о том, что отдельные пары признаков ведут себя в наследовании независимо. В этом сущность третьего закона Менделя — закона независимого наследования признаков, или независимого комбинирования генов.
Он формулируется так: каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга.
Закон независимого комбинирования генов составляет основу комбинативной изменчивости (см. Рис. 4), наблюдаемой при скрещивании у всех живых организмов. Отметим также, что в отличие от первого закона Менделя, который справедлив всегда, второй закон действителен только для генов, локализованных в разных парах гомологичных хромосом. Это обусловлено тем, что негомологичные хромосомы комбинируются в клетке независимо друг от друга, что было доказано не только при изучении характера наследования признаков, но и прямым цитологическим методом. Поведение хромосом при дигибридном скрещивании показано на рис. 4.
Цитологические основы дигибридного скрещивания. Как известно, в профазе I мейоза гомологичные хромосомы конъюгируют, а в анафазе одна из гомологичных хромосом отходит к одному полюсу клетки, а другая — к другому. При расхождении к разным полюсам негомологичные хромосомы комбинируются свободно и независимо друг от друга. При оплодотворении в зиготе восстанавливается диплоидный набор хромосом и гомологичные хромосомы, оказавшиеся в процессе мейоза в разных половых клетках родителей, соединяются вновь.
Предположим, что каждая хромосома содержит только один ген. Палочковидные хромосомы несут аллель A или а, сферические —В или b, т. е. эти две пары аллелей находятся в негомологичных хромосомах (см. рис. 4).
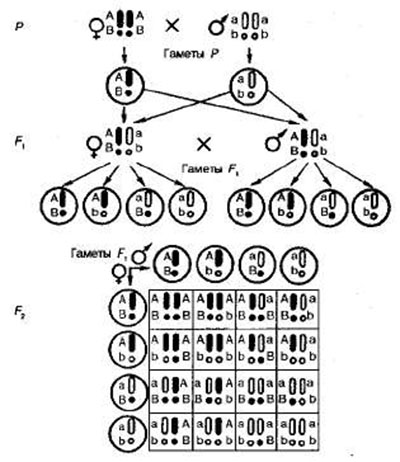
Рис 4. Цитологические основы расщепления признаков при дигибридном скрещивании.
Гомозиготные родители (ААВВ и aabb) формируют только один тип гамет с доминантными (АВ) или с рецессивными (ab) аллелями. При слиянии таких гамет образуется единообразное первое поколение гибридов — гибрид дигетерозиготен (АаВb), но так как у него присутствуют гены А и B, то по фенотипу он сходен с одним из родителей.
В тех случаях, когда необходимо указать, что те или иные гены находятся в гомологичных хромосомах, в генетических формулах зигот хромосомы принято изображать в виде двух черточек или одной с указанием обоих аллелей гена. Формула дигетерозиготы может быть записана так:. Поскольку гаметы содержат только по одной из гомологичных хромосом и соответственно по одному аллелю каждого гена, то их формулы могут быть записаны так: и т. д.
В дальнейшем у гибридных организмов по причине случайности расхождения отцовских и материнских хромосом каждой пары в процессе мейоза ген А может попасть в одну гамету с геном В или с геном Ь. Точно так же ген а может оказаться в одной гамете с геном В или с геном b. Поэтому гибриды образуют четыре типа гамет: Образование всех четырех типов гамет равновероятно, т. е. все они образуются в равных количествах. Свободное сочетание таких гамет в процессах оплодотворения заканчивается образованием 16 типов зигот, а значит, и потомков (см. рис. 4).
Они распадаются на четыре фенотипических класса: доминантные по обоим признакам — 9 частей, доминантные по первому и рецессивные по второму признаку — 3 части, рецессивные по первому и доминантные по второму — 3 части, рецессивные по обоим признакам — 1 часть. Генотипических классов 9: 1AABB, 2ААВb, 1AAbb, 1Aabb, 4AaBb, 2AaBB, 1aaBB, 2aaBb, 1aabb.
Полигибридное скрещивание. Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании, т. е. когда родители различаются по аллелям трех и более генов, а в F1 образуются три- и по дигетерозиготы. Соотношение генотипических и фенотипических классов в F2 три- и полигибридных скрещиваний, а также число типов гамет (и число фенотипов) у гибридов F1 определяются простыми формулами: при моногибридном скрещивании число типов гамет равно 2, при дигибридном 4 (22), а при полигибридном — 2n; число генотипов равно соответственно 3,9 (32) и 3n.
Опираясь на независимость наследования разных пар аллелей, можно также любые сложные расщепления представить как произведение от соответствующего числа независимых моногибридиых скрещиванй. Общая формула определения фенотипических классов при полигибридном скрещивании имеет вид (3:1)n, где n равно числу пар признаков, по которым идет расщепление. Для моногибрида эта формула соответственно имеет вид (3:1); дигибрида — 9:3:3:1 или (3:1)2;тригибрида — (3:1)3. Расщепление по генотипу имеет вид (1:2; 1)n, где n — число расщепляющихся пар аллелей.
Известно, что каждый организм гетерозиготен по многим генам. Если предположить, что человек, у которого отдельные пары хромосом содержат не одну, а сотни пар аллелей, гетерозиготен хотя бы по 20 генам, то число типов гамет у такой полигетерозиготы составит 220 = 1 048 576. Эта цифра дает определенное представление о потенциальных возможностях комбинативной изменчивости. Поэтому каждый человек обладает неповторимой индивидуальностью. На Земле нет двух людей, совершенно одинаковых по наследственности, за исключением однояйцевых близнецов.
Таким образом, третий закон Менделя (закон независимого наследования признаков) еще раз демонстрирует дискретный характер генетического материала. Это проявляется в независимом комбинировании аллелей разных генов и в их независимом действии — фенотипическом выражении.
Дискретность гена определяется тем, что он контролирует присутствие или отсутствие отдельной биохимической реакции, от которой зависит развитие или подавление определенного признака организма. Очевидно, если несколько генов определяют какое-либо одно свойство или один признак (форма гребня у кур, окраска глаз у дрозофилы, длина колоса у пшеницы и т. д.), они должны взаимодействовать между собой. Отсюда следует, что понятие «наследование признаков» употребляется, скорее всего, как образное выражение, поскольку в действительности наследуются не сами признаки, а гены. Признаки формируются в ходе индивидуального развития организма, обусловливаются генотипом и влиянием внешней среды.
Домашних животных человек разводит в течение тысячелетий путем скрещивания, отбора и подбора с целью совершенствования существующих и создания новых пород. Отобранные для разведения собаки должны обладать желательным типом конституции, уравновешенным типом поведения, хорошими слухом, зрением, обонянием, способностью к дрессировке и консервативной наследственностью.
В зоотехнии различают три основных метода разведения: чистопородное разведение, скрещивание и гибридизацию.
В клубах служебного, охотничьего и декоративного собаководства применяют метод чистопородного разведения, при котором кобеля одной породы спаривают с сукой этой же породы. Различают чистопородное разведение по генеалогическим заводским линиям и по семействам.
Чистопородный метод разведения позволяет закрепить отличительные черты породы и быстро добиться однородности потомства. Этим методом добиваются постепенного совершенствования породы и, как следствие этого, улучшения основных ее качеств. В то же время необходимо опасаться закрепления отрицательных качеств и понижения жизнеспособности потомства. Чистопородное разведение бывает двух типов: родственное (инбридинг) и неродственное (аутбридинг).
Родственное разведение — инбридинг — это скрещивание между собой животных, находящихся в той или иной степени родства. Инбридинг является одним из сильных зоотехнических методов воздействия на продуктивные и рабочие качества животных. Он может оказывать на них как положительное, так и отрицательное влияние.
Применение инбридинга способствует концентрации наследственных задатков, в результате чего получают животных, гомозиготных по ряду признаков, что позволяет быстро закреплять ценные свойства и создавать однородное поголовье. В то же время неумелое и бессистемное применение инбридинга приводит к обеднению генотипа и концентрации нежелательной наследственности, т. е. к ослаблению конституции, снижению продуктивности и резистентности у потомства, а иногда и к появлению уродств в последующих поколениях.
Для инбридинга необходимо отбирать абсолютно здоровых животных, правильно содержащихся, имеющих желательный тип конституции и хорошие рабочие качества. Инбридинг при разведении собак применяется в тех случаях, когда хотят быстро закрепить у потомков качества выдающегося производителя или свойства, присущие линии или семейству.
В собаководстве различают тесное, близкое, умеренное и отдаленное родственное разведение (инбридинг). При тесном инбридинге спаривают отца с дочерью, мать с сыном. Такое спаривание относят к I степени родства. Близкородственное разведение осуществляется при спаривании дедов с внуками, братьев с сестрами, прадедов с правнуками и дядей с племянниками (II и III степени родства).
При умеренном родственном разведении спаривают животных, состоящих в IV, V, VI степенях родства. Отдаленным родственным разведением считают такое, при котором спаривают животных, находящихся в более отдаленных степенях родства.
В основном метод родственного разведения применяют в случаях, когда животные состоят в умеренных и отдаленных степенях родства. Близкородственное разведение может быть допущено как исключение в племенных хозяйствах и питомниках с целью закрепления желательных признаков. Тесный инбридинг при разведении пользовательных собак недопустим.
Степень инбридинга обозначается римскими цифрами. Если общий предок встречается в первом ряду родословной карточки, его обозначают цифрой I, во втором — II и т. д., причем повторяющийся предок со стороны отца пишется первым, а со стороны матери — вторым после тире, например: Чемпион Ингул I — II, III (Ингул в родословной матери стоит как во втором, так и в третьем ряду предков). Если повторяется пара предков, налицо комплексный инбридинг, закрепляющий свойства удачно подобранной пары.
В результате чистопородного родственного разведения образуются родственные группы собак, имеющие выдающегося общего предка, линии. Различают кровные и заводские линии.
Кровные линии объединяют всех собак, происходящих от одного родоначальника, независимо от их типа и качеств. Животные одной кровной линии бывают разнообразны по своим рабочим и экстерьерным качествам, и поэтому со всей кровной линией племенную работу не ведут.
Заводская генеалогическая линия выделяется из кровной. К ней принадлежат те животные, которые соответствуют основному типу кровной линии и обладают ценными качествами ее выдающегося производителя.
Каждая порода собак имеет свои выдающиеся линии и называется именем своего производителя, например, у восточноевропейских овчарок была линия Абрека и линия Грея; у ротвейлеров — линия Карена и линия Брикса и т. п. В каждой породе постепенно выделяются новые линии. Обычно в роли родоначальника линии выступает выдающийся кобель, дающий большое количество потомков.
Суки многоплодны, поэтому в течение нескольких лет способны дать большое количество щенков от одного или разных кобелей-производителей. Группу потомков от одной выдающейся суки называют семейством. Линии и семейства очень тесно связаны между собой. В своем развитии линия опирается на одно или несколько семейств.
Ограниченность генофонда заводских линий и родство всего поголовья обусловливают в отдельных случаях родственное скрещивание потомства от производителя с наиболее желательными породными признаками, т. е. проводить разведение «в себе». Такое разведение в отдаленных рядах предков считают положительным, так как оно способствует консолидации породы, закреплению ее отличительных признаков.
Чистопородное разведение, при котором кобелей одной линии спаривают с суками другой линии, называют межлинейным. Такое разведение способствует обогащению линии полезными качествами и позволяет избежать тесного и близкого инбридинга, повышает жизнеспособность животных.
Одним из методов разведения собак является межпородное скрещивание — аутбридинг. Оно используется в основном при выведении новых пород собак.
Различают промышленное, или пользовательное, скрещивание, вводное скрещивание (прилитие крови), поглотительное, воспроизводительное скрещивание и межвидовую гибридизацию.
Промышленное скрещивание используется для получения пользовательных, т.е. неплеменных, животных. При промышленном скрещивании спаривают собак двух разных пород, дополняющих друг друга по каким-либо желательным качествам. При этом в работе используются только помеси первого поколения.
Эффект скрещивания основан на явлении гетерозиса, заключающегося в том, что у помесей (гибридов) первого поколения желательные признаки и свойства проявляются в большей степени, чем у их родителей. Во всех случаях, когда молодым собакам создают необходимые условия кормления и содержания, они намного превосходят своих родителей по выносливости, величине, злобности и другим желаемым качествам.
Вводное скрещивание применяется тогда, когда в породе в дополнение к имеющимся у нее признакам желают добавить другие, недостающие или утраченные ею. Для этого подбирают кобеля породы, близкой к улучшаемой по основным качествам, но обладающей ярко выраженным желательным признаком, и спаривают с ним несколько сук породы, которую хотят улучшить. Полученных в результате вводного скрещивания помесных потомков спаривают в дальнейшей с собаками улучшаемой (основной) породы.
В практике собаководства вводное скрещивание успешно применяется. Например, первым представителем эрдельтерьеров приливали кровь бультерьера. При вводном скрещивании могут быть использованы две, три и более породы. Так, при выведении новой породы черный терьер, были использованы собаки трех пород: эрдельтерьер, ризеншнауцер и ротвейлер. Затем путем отбора и подбора уже при чистопородном разведении из породной группы была создана порода черный терьер, успешно применяемая для несения караульной службы.
Поглотительное скрещивание применяют тогда, когда хотят поглотить (вытеснить) какую-либо местную породу, не отвечающую предъявляемым требованиям, другой, обладающей всеми необходимыми качествами. Помесные суки, полученные в результате спаривания сук улучшаемой породы с производителями улучшающей снова спариваются с производителями улучшающей породы и т.д., до тех пор пока полученное поколение не приблизиться по качеству к улучшающей породе.
Поглотительное скрещивание в собаководстве особого применения не нашло.
Основной целью воспроизводительного скрещивания является выведение новой породы, соединяющей в себе лучшие качества нескольких взятых пород. Почти все современные культурные породы собак получены в результате воспроизводительного скрещивания двух или нескольких пород. Например, южнорусская овчарка произошла от астурийской овчарки, местных степных борзых и азиатских овчарок, доберман — от немецкого пинчера, ротвейлера и французской овчарки босерон и т. п.
Генетическая сущность инбридинга и его значение для селекции. Влияние инбридинга на генетическую структуру популяции.
Инбридинг – один из методов разведения животных. Селекционерам важно знать особенности этого метода, чтобы правильно использовать его в племенной работе. Незнание особенностей инбридинга, его влияния на результаты разведения может привести к неправильному его применению, а значит, к ухудшению качества поголовья.
Инбридинг – это система спаривания животных, находящихся в родстве. Родственные спаривания необходимы для того, чтобы генетически закрепить признаки, то есть добиться гомозиготности по генам, их определяющим. Некоторая часть лучшего в мире поголовья собак и других домашних животных была получена путем инбридинга. Но, применяя инбридинг без достаточных знаний, можно погубить породу за несколько генераций.
Краткая история инбридинга.
Учение об инбридинге возникло давно. В прошлом родственное спаривание применялось в животноводстве бессознательно, стихийно. В древние времена, когда люди обнаружили вредные действия кровосмешения, существовали строгие законы, запрещающие родственное спаривание. У арабов уже в 18 веке накопилось много фактов отрицательного влияния на потомство племенных лошадей. В период развития капитализма начался процесс породообразования во многих странах. Создание новых пород, отвечающих требованиям рынка, осуществлялось с широким использованием инбридинга. В то время были получены множество известных пород сельхоз.животных, в том числе орловская порода лошадей. Известный селекционер прошлого Р. Беквелл основой создания новых пород считал использование инбридинга любых степеней с целью закрепления в потомстве выдающихся качеств родоначальника.
Но очень широкое, порой непродуманное, без всякой цели применение тесного инбридинга скоро обернулось для многих заводчиков большой бедой. Бессистемный тесный инбридинг привел к снижению продуктивности и плодовитости, крепости конституции животных. В результате этого взгляды на инбридинг начали меняться. К этому методу спаривания стали относиться осторожно. В ту пору не существовало теорий, объясняющих причины вредных последствий родственного спаривания, но инбридинга стали избегать. Интерес к инбридингу проявился вновь только к концу 19 века. Анализ племенных книг, изучение истории создания многих заводских пород показали положительную роль инбридинга в породообразовании. С начала 20 века вопрос о роли и значении инбридинга, его биологической сущности начинают решать генетики.
Большинство пород собак изначально были получены в результате инбридинга. Как правило, родоначальник породы — это одна или несколько особей, которых спаривали между собой в близком родстве, дабы получить новое поколение животных. Этот метод используется для стабилизации типа и уменьшения внутрипородных вариаций. Он дает не только чисто внешнюю схожесть собак, но приводит и к концентрации генофонда. Инбридинг не может внести никаких новых пороков развития, кроме тех, которые были заложены в генотипе собак. Этот метод как раз и позволяет вскрыть имеющиеся после ауткросса (неродственного скрещивания), но не проявившееся внешне аномальные гены. Если вы действительно хотите знать пороки и недостатки вашей линии, используйте на протяжении пары генераций инбридинг, и все тайное станет явью. Именно поэтому средние заводчики избегают его, обвиняя других в воспроизводстве пороков и недостатков. Но виной тому вовсе не инбридинг, а аномальные гены, которые находились у родителей в скрытой форме. Метод позволил им проявиться внешне, а не породил новые нарушения развития.
Начинающим собаководам, которые не знают собак, перечисленных в родословной по крайней мере за четыре поколения назад, нецелесообразно пытаться применять инбридинг. Рискованные эксперименты очень дорого стоят и могут принести большие огорчения. Бывает, что инбридинг выявляет недостатки предков, отстоящих на несколько поколений назад.
Во многих породах собак при разведении сегодня применяется инбридинг. Запись проводят римскими цифрами, начиная с материнской стороны родословной. Степень родства может быть разной: очень тесный инбридинг (кровосмешение) – сестра х брат, мать х сын, дочь х отец (II – II; II – I; I – II) ; близкий инбридинг (тесное родственное спаривание) – бабушка х внук, внучка х дед (III – II; II – III); умеренный инбридинг – общий предок встречается в III – IV поколениях; отдаленный инбридинг – общий предок встречается в V поколениях. Если общий предок отмечается в родословной дальше V поколения, животные считаются неродственными. Степень инбридинга зависит как от места общего предка в родословной, так и от сходства животных по типу, породности.
Основная цель инбридинга – сохранение конкретных наследственных особенностей того или иного выдающегося предка. Инбридинг производится на выдающегося по своим экстерьерным качествам производителя (реже — на пару производителей), который, в идеале, должен обладать высокой препотентностью по комплексу значимых признаков. Кроме того, этот производитель не должен быть носителем рецессивно наследуемых аномалий. В результате инбридинга на выдающегося производителя через его лучших потомков, которые унаследовали тип общего предка и его главные достоинства, происходит накопление и наследственное закрепление ценных признаков и повышение способности передавать их потомству. У потомков происходит накопление доминантных генов, отвечающих за выдающиеся признаки. При условии отбраковки и оставлении только лучших особей, которые наиболее напоминают того предка, на которого сделан инбридинг, эти особи, будучи доминантными по многим признакам, закрепленных в них, будут передавать их своим потомкам. Но нужно учитывать и знать рецессивные гены, которые несет общий предок, так как наряду с достоинствами потомки наследуют и его недостатки, которые могут быть закреплены в рецессивных генах.
Однако длительное применение инбридинга, особенно при спаривании близких родственников, у полученного потомства может проявляться инбредная депрессия — это снижение иммунных свойств организма, половой потенции, плодовитости, появление нарушений в период беременности, отставание в росте у животных — действительно может наблюдаться при длительных (в течение нескольких поколений) инбридингах. Связана она не со специфическими генами заболеваний или аномалий, а с накоплением аллелей комплексного действия, снижающих жизнеспособность.
Велика роль инбридинга при разведении по линиям. Такое разведение, по признанию многих ученых, — высшая форма племенной работы при чистопородном разведении. Линия представляет собой группу высокоценных по продуктивным и племенным качествам животных, происходящих от одного выдающегося родоначальника и сходных с ним по конституции и продуктивности. Главная цель применения инбридинга при этом — закрепление наследственности родоначальника линии и повышение гомозиготности потомства.
Вернемся к тому, что история кинологии знает немало примеров, когда при помощи этого метода были выведены и сохранены прекрасные породы собак, получены выдающиеся производители и чемпионы. Возьмем, к примеру, наиболее распространенную породу собак – немецкую овчарку. На ранних этапах формирования немецкой овчарки основатель породы Макс фон Штефаниц применял близкий и даже тесный инбридинг на лучших производителей (основателей породы) – Хоранда ф. Графрат, Гектора ф. Швабен, Хайнца ф. Штаркенбург. Спустя десятилетие ряд чемпионов также были получены при помощи близкородственного разведения. Это способствовало закреплению в популяции лучших качеств выдающихся производителей и быстрому формированию однотипности породы. Когда была создана достаточно однородная популяция немецкой овчарки, у заводчиков отпала необходимость в близких инбридингах. Вместо них стали широко использоваться умеренные и отдаленные инбридинги на выдающихся животных. Так, например, родоначальники всех современных генеалогических линий в породе получены при помощи умеренного и отдаленного инбридинга на лучших собак того времени. В настоящее время заводчики Германии редко используют близкий инбридинг. В этом нет необходимости, так как порода, несмотря на свою многочисленность, достаточно однородна в типе, анатомических особенностях, и находится на верном пути прогресса.
Подводя итог сказанному, можно сделать вывод, что инбридинг как метод разведения имеет большое значение в породообразовании и разведении в общем. Но при всем при этом, неумелое применение инбридинга может привести к отрицательным последствиям, негативно отразиться для породы. Применение инбридинга должно носить целенаправленный характер и осуществляться только на высококлассных предков, тип и качество которых планируется закрепить в потомстве. Самое главное, их влияние на популяцию должно быть актуально на сегодняшний день.
Термин «популяция» происходит от латинского populus – население. Долгое время (начиная с конца XVIII в.) популяцией называли (а часто называют и сейчас) любую группировку организмов, обитающих на определенной территории.
В 1903 г. датский генетик Вильгельм Людвиг Иоганнсен впервые употребил термин «популяция» для обозначения группы особей, неоднородной в генетическом отношении.
С точки зрения генетики, популяция – это генетическая система, обладающая исторически сложившейся генетической структурой. Основные положения популяционной генетики сложились на основании изучения природных и модельных популяций высших раздельнополых животных (моллюсков, насекомых, позвоночных), которые воспроизводят себя с помощью нормального полового размножения – амфимиксиса, или объединения женских и мужских гамет. В таких случаях группировка особей, способных скрещиваться между собой и производить полноценное (т.е. жизнеспособное и плодовитое) потомство, называется генетической, или менделевской популяцией. В свою очередь, потомки, достигшие половозрелости, также должны скрещиваться между собой и производить полноценное потомство, то есть популяция должна существовать длительное число поколений.
Таким образом, с точки зрения генетики, популяция представляет собой множество особей, объединенных достаточно высокой степенью родства.
Генетическая структура популяций.
Каждая популяция обладает собственной генетической структурой. Генетическая структура популяций определяется исходным соотношением аллелей, естественным отбором и элементарными эволюционными факторами (мутационный процесс и давление мутаций, изоляция, популяционные волны, генетико-автоматические процессы, эффект основателя, миграции и др.). Для описания генетической структуры популяций используются понятия «аллелофонд» и «генофонд».
Аллелофонд. Аллелофонд популяции – это совокупность аллелей в популяции. Если рассматриваются два аллеля одного гена: А и а, то структура аллелофонда описывается уравнением: pA + qa = 1. В этом уравнении символом pA обозначается относительная частота аллеля А, символом qa – относительная частота аллеля а.
Популяции, в которых структура аллелофонда остается относительно постоянной в течение длительного времени, называются стационарными.
Если рассматриваются три аллеля одного гена: а1, а2,, а3, то структура аллелофонда описывается уравнением: p а1 + q а2 + r а3 = 1. В этом уравнении символами p, q, r обозначаются соответствующие частоты аллелей.
Если рассматриваются несколько аллелей нескольких генов (a, b, c), то структура аллелофонда описывается системой уравнений:
p1 a1 + p2 a2 + p3 a3 + … + pi ai = 1
q1 b1 + q2 b2 + q3 b3 + … + qi bi = 1
r1 c1 + r2 c2 + r3 c3 + … + ri ci = 1
В этих уравнениях символами pi, qi, ri обозначены относительные частоты аллелей разных генов. Однако в простейших случаях рассматриваются только моногенные диаллельные системы, например: А–а. В популяции с общей численностью особей Nобщ и известной численностью особей с генотипами АА, Аа, аа относительные частоты аллелей рассчитываются по формулам:
|
p (A) = |
2 Í N (AA) + N (Aa) |
|
2 Í N общ. |
|
q (a) = |
2 Í N (aa) + N (Aa) |
|
2 Í N общ. |
|
или q (a) = 1 – р (А) |
|
|
|
Генофонд. Термин генофонд употребляется в разных значениях. Основоположник учения о генофонде и геногеографии Александр Сергеевич Серебровский называл генофондом «совокупность всех генов данного вида…, чтобы подчеркнуть мысль о том, что в лице генофонда мы имеем такие же национальные богатства, как и в лице наших запасов угля, скрытых в наших недрах» (1928). Однако это выражение в настоящее время используется для определения генетического потенциала, а генофондом называют совокупность всех генотипов в популяции.
При изучении природных популяций часто приходится сталкиваться с полным доминированием: фенотипы гомозигот АА и гетерозигот Аа неразличимы. Кроме того, в природе широко распространено полигенное определение признаков, причем типы взаимодействия неаллельных генов (комплементарность, эпистаз, полимерия) не всегда известны. Поэтому на практике часто изучают не генофонд, а фенофонд популяций, то есть соотношение фенотипов. В настоящее время развивается раздел генетики популяций, который называется фенетика популяций.
Закон Харди–Вайнберга – основной закон популяционной генетики
Структура генофонда в панмиктической стационарной популяции описывается основным законом популяционной генетики – законом Харди-Вайнберга, который гласит, что в идеальной популяции существует постоянное соотношение относительных частот аллелей и генотипов, которое описывается уравнением:
(p A + q a)2 = р2 АА + 2∙р∙q Aa + q2 aa = 1
Если известны относительные частоты аллелей p и q и общая численность популяции Nобщ, то можно рассчитать ожидаемую, или расчетную абсолютную частоту (то есть численность особей) каждого генотипа. Для этого каждый член уравнения нужно умножить на Nобщ:
p2 AA · Nобщ + 2·p·q Aa · Nобщ + q2 aa · Nобщ = Nобщ
В данном уравнении:
p2 AA · Nобщ – ожидаемая абсолютная частота (численность) доминантных гомозигот АА
2·p·q Aa · Nобщ – ожидаемая абсолютная частота (численность) гетерозигот Аа
q2 aa · Nобщ – ожидаемая абсолютная частота (численность) рецессивных гомозигот аа
Неслучайное в отношении фенотипа скрещивание происходит в популяциях, где скрещивающиеся особи находятся в большем или меньшем родстве друг к другу. Такие скрещивания называются инбридингом в отличие от случайного аутбридинга. Ни инбридинг, ни аутбридинг, сами по себе, не приводят к изменению частот аллелей, но оба приводят к перераспределению аллелей в генотипах. В инбредной популяции частота гомозигот повышена, а частота гетерозигот понижена относительно соотношений Харди-Вайнберга. При аутбридинге наблюдается обратное соотношение: частота гетерозигот повышена, а гомозигот — понижена по сравнению с пропорциями при случайном скрещивании. Конечно, внутри популяции, не имеющей структуры, эффект перекрестного оплодотворения (аутбридинга) не может быть велик. Только когда особи предпочтительно скрещиваются с особями из другой популяции (или субпопуляции) наблюдается значительное влияние перекрестного оплодотворения на частоты генотипов.
Следует отметить несколько других общих закономерностей. Во-первых, генотипические изменения, обусловленные инбридингом, проявляются во всех локусах генома. Заметьте, что на все локусы влияет генный поток, а отбор или мутация действуют только на единственный локус (или на тесно сцепленные с ним локусы). Во-вторых, влияние на частоты генотипов может быть совершенно незначительным, если система скрещивания изменится. Например, высокая частота гомозигот, являющаяся результатом самооплодотворения, может быть полностью уничтожена одним поколением случайного скрещивания. Наконец, инбридинг и генетический дрейф, в общем, сходно влияют на гетерозиготность, но результаты их действия различны. Инбридинг (в большой популяции) может привести к понижению гетерозиготности без изменения частот аллелей данного локуса в популяции. С другой стороны, генетический дрейф может вызвать изменение частоты аллелей, но не приведет к понижению гетерозиготности в популяции. Только при рассмотрении усредненного влияния генетического дрейфа на данный локус во многих популяциях, или на многие локусы в данной популяции, можно выявить зависимость уровня гетерозиготности от генетического дрейфа.
Мутационная изменчивость как одна из форм наследственной изменчивости.
Мутационной называется изменчивость самого генотипа. Мутации — это внезапные наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
Существование наследственных изменений было известно Дарвину. Вся его теория эволюции вытекает из учения о естественном отборе наследственных изменений. Наследственная изменчивость — необходимая предпосылка естественного и искусственного отбора. Однако во времена Дарвина еще отсутствовали опытные данные по наследственности и законы наследования не были известны. Это не давало возможности строго различать разные формы изменчивости в зависимости от наследования.
Понятие мутаций было введено в науку голландским ботаником-де Фризом. У растения ослинник (энотера) он наблюдал появление резких скачкообразных отклонений от типичной формы растения, причем эти отклонения оказались наследственными. Дальнейшие исследования на различных объектах — растениях, животных, микроорганизмах — показали, что явление наследственной (мутационной) изменчивости свойственно всем организмам. Мутации затрагивают разнообразные стороны строения и функции организма. Например, у дрозофилы известны мутационные изменения формы крыльев (вплоть до полного их исчезновения), окраски тела, развития щетинок на теле, формы глаз, их окраски (красные, желтые, белые, вишнёвого цвета и т. п.), а также многих физиологических признаков (продолжительность жизни, плодовитость, стойкость к разным повреждающим воздействиям и т. п.). Первоначальные представления-де Фриза о том, что мутации всегда крупные наследственные изменения, дальнейшими исследованиями не подтвердились. Наряду с резкими отклонениями гораздо чаще встречаются небольшие мутации, лишь немногим отличающиеся от исходных форм. Тем не менее, указанный еще-де Фризом признак мутаций — их скачкообразный характер и наследственность — остается в силе. Мутации совершаются в различных направлениях, и обычно они не являются приспособительными, полезными для организма изменениями.
Существуют и такие наследственные изменения, которые в гомозиготном состоянии вызывают гибель (такие мутации называются летальными).
Частота и причины мутаций
Как часто происходят мутации? Каковы причины их возникновения? Прежде чем ответить на этот вопрос, нужно иметь в виду, что учет возникающих мутаций представляет очень большие трудности. Большинство мутаций рецессивны. Они возникают в генах, локализованных в хромосомах половых клеток. Гамета, несущая вновь возникшую рецессивную мутацию, при оплодотворении обычно соединяется с гаметой, которая такой же мутации не несет. Поэтому вновь возникшая рецессивная мутация фенотипически не проявится. Однако в последующих поколениях она будет размножаться вместе с несущей ее хромосомой, и распространяться среди особей данного вида. Лишь когда соединятся две гаметы, несущие одну и ту же рецессивную мутацию, она проявится фенотипически.
Исследования показали, что в природных условиях мутация каждого отдельно взятого гена происходит очень редко. На первый взгляд может возникнуть представление, что такая малая изменчивость гена не может дать достаточного материала наследственной изменчивости для естественного отбора. На самом деле это не так. У организма имеется несколько тысяч генов, так что общее число мутаций оказывается значительным. Для той же дрозофилы, например, высчитано, что около 5% ее гамет несут какую-нибудь мутацию. Прямые исследования распространения мутаций в природных популяциях дрозофилы, проведенные в разных географических зонах, показали, что они «насыщены» разнообразными мутациями, большинство которых, однако, в силу рецессивности не проявляется видимо. Значительная стойкость гена имеет большое биологическое значение. Действительно, если бы гены легко и часто изменялись, то существование видов стало бы невозможным, ибо в каждом поколении организмы превращались бы в нечто совершенно новое, не похожее на родителей. Относительная стойкость видов — важное условие приспособления организма к окружающей среде.
Способность к мутированию — одно из основных свойств гена. Разумеется, каждая отдельная мутация вызывается какой-то причиной. Однако в большинстве случаев эти причины остаются нам неизвестными. Мутации связаны с изменениями во внешней среде. Это убедительно доказывается тем, что различными внешними факторами удается резко повысить число возникающих мутаций. Особенно эффективно действующими факторами экспериментального получения мутаций оказываются такие, которые влияют на нуклеиновые кислоты. Это вполне понятно, так как материальной основой генов служит ДНК.
Впервые в опыте резкое повышение числа возникающих наследственных изменений было получено действием лучей Рентгена. Под влиянием рентгенизации число получаемых мутаций удалось повысить в 150 раз и даже более. С тех пор экспериментальное получение мутаций было осуществлено на самых различных организмах — от бактерий и вирусов до млекопитающих и цветковых растений. Кроме лучей Рентгена и других форм ионизирующей радиации, мутации могут быть вызваны самыми различными химическими и физическими воздействиями: температурой, изменением газового режима, влажности и т. п. Любые изменения, затрагивающие процессы обмена веществ, оказывают свое влияние и на мутационный процесс. Результаты исследований по экспериментальному получению мутаций показали, что в основном дело сводится к увеличению их частоты. Экспериментально вызываемые наследственные уклонения совершаются в различных направлениях, так же как и естественный процесс мутационной изменчивости. Лишь в самое последнее время намечаются некоторые пути воздействия на направление мутаций. Эти новые возможности базируются на глубоком проникновении в механизм процесса синтеза нуклеиновых кислот.
Экспериментальное получение мутаций имеет и большое практическое значение, так как резко повышает наследственную изменчивость, давая, таким образом, материал для отбора.
Важная закономерность была установлена Н.И. Вавиловым. Она известна под именем закона гомологических рядов наследственной изменчивости. Сущность этого закона сводится к тому, что виды и роды, генетически близкие (т. е. связанные друг с другом единством происхождения), характеризуются сходными рядами наследственной изменчивости. Зная наследственные изменения у одного вида, можно предвидеть нахождение сходных изменений у родственных видов и родов.
Основные типы мутаций и их классификаций
Генные (точковые) мутации.
Затрагивают, как правило, один или несколько нуклеотидов, при этом один нуклеотид может превратиться в другой, может выпасть (делеция), продублироваться, а группа нуклеотидов может развернутся на 180 градусов. Генные мутации приводят к изменению аминокислотной последовательности белка. Наиболее вероятная мутация генов происходит при спаривании тесно связанных организмов, которые унаследовали мутантный ген у общего предка. Интересно, что значимость нуклеотидных мутаций внутри кодона неравнозначна: замена первого и второго нуклеотида всегда приводит к изменению аминокислоты, третий же обычно не приводит к замене белка. К примеру, «Молчащая мутация»- изменение нуклеотидной последовательности, которая приводит к образованию схожего кодона, в результате аминокислотная последовательность белка не меняется.
Геномные мутации.
Главная отличительная черта геномных мутаций связана с нарушением числа хромосом в кариотипе. Эти мутации так же подразделяются на два вида: полиплоидные анеуплоидные.
Полиплоидные мутации ведут к изменению хромосом в кариотипе, которое кратно гаплоидному набору хромосом. Этот синдром впервые был лишь обнаружен в 60-ых годах. Вообще полиплодия характерна в основном для человека, а среди животных встречается крайне редко. При полиплоидии число хромосом в клетке насчитывается по 69 (триплодие), а иногда и по 92 (тетраплодие) хромосомы. Такое изменение ведет практически к 100% смерти зародыша. Триплодие имеет не только многочисленные пороки, но и приводит к потере жизнеспособности. Тетраплодие встречается еще реже, но так же зачастую приводит к летальному исходу.
Анеуплоидные мутации приводят к изменению числа хромосом в кариотипе, некратные — гаплоидному набору. В результате такой мутации возникают особи с аномальным числом хромосом. Как и триплодия, анеуплодия часто приводит к смерти еще на ранних этапах развития зародыша. Причиной же таких последствий является утрата целой группы сцепления генов в кариотипе.
В целом же, механизм возникновения геномных мутаций связан с патологией нарушения нормального расхождения хромосом в мейозе, в результате чего образуются аномальные гаметы, что и ведет к мутации. Изменения в организме связаны с присутствием генетически разнородных клеток. Такой процесс называется мозаицизм.
Геномные мутации одни из самых страшных.
Хромосомные мутации.
Хромосомные мутации приводят к изменению числа, размеров и организации хромосом, поэтому их иногда называют хромосомными перестройками. Хромосомные перестройки делятся на внутри- и межхромосомные. К внутрехромосмным относятся:
Дубликация — один из участков хромосомы представлен более одного раза.
Делеция — утрачивается внутренний участок хромосомы.
Инверсия — повороты участка хромосомы на 180 градусов.
Межхромосомные перестройки (их еще называют транслокации) делятся на:
Реципрокные — обмен участками негомологичных хромосом.
Нереципрокные — изменение положения участка хромосомы.
Дицентрические — слияние фрагментов негомологичных хромосом.
Центрические — слияние центромер негомологичных хромосом.
Классификация мутаций
Трудности определения понятий «мутация» лучше всего иллюстрирует классификация ее типов.
Существует несколько принципов такой классификации:
A). По характеру изменения генома:
1. Геномные мутации — изменение числа хромосом.
Геномные мутации — это мутации, которые приводят к добавлению либо утрате одной, нескольких или полного гаплоидного набора хромосом. Разные виды геномных мутаций называют гетероплоидией и полиплоидией.
Геномные мутации связаны с изменением числа хромосом. Например, у растений довольно часто обнаруживается явление полиплоидии — кратного изменения числа хромосом. У полиплоидных организмов гаплоидный набор хромосом n в клетках повторяется не 2, как у диплоидов, а значительно большее число раз (3n, 4п, 5п и до 12n). Полиплоидия — следствие нарушения хода митоза или мейоза: при разрушении веретена деления удвоившиеся хромосомы не расходятся, а остаются внутри неразделившейся клетки. В результате возникают гаметы с числом хромосом 2n. При слиянии такой гаметы с нормальной (n) потомок будет иметь тройной набор хромосом. Если геномная мутация происходит не в половых, а в соматических клетках, то в организме возникают клоны (линии) полиплоидных клеток. Нередко темпы деления этих клеток опережают темпы деления нормальных диплоидных клеток (2n). В этом случае быстро делящаяся линия полиплоидных клеток образует злокачественную опухоль. Если она не будет удалена или разрушена, то за счет быстрого деления полиплоидные клетки вытеснят нормальные. Так развиваются многие формы рака. Разрушение митотического веретена может быть вызвано радиацией, действием ряда химических веществ-мутагенов.
Геномные мутации в животном и растительном мире очень многообразны.
2. Хромосомные мутации, или хромосомные перестройки, — изменение структуры хромосом.
Хромосомные мутации — это перестройки хромосом. Участки хромосом могут изменить свое положение, потеряться или удвоиться.
Хромосомные мутации — это мутации, нарушающие существующие группы сцепления или приводящие к возникновению новых групп сцепления. Такое определение указывает на способ, которым эти мутации в первую очередь обнаруживаются. Согласно другому определению, хромосомные мутации — это мутации, обусловленные перестройками хромосом. Хромосомные перестройки бывают разных типов. Пожалуй, наиболее распространенная — рекомбинация, или кроссинговер, при котором происходит обмен гомологичными участками хромосом. Другие типы перестроек хромосом — это транслокации, инверсии, делеции и дупликации.
Разнообразны варианты изменения морфологии хромосом. Различают следующие ХП: — Реципрокные транслокации — обмен участками хромосом. — Робертсоновские транслокации — слияние двух акроцентрических хромосом в одну двуплечую хромосому. — Парацентрическая инверсия — изменение порядка генов на обратный в пределах участка, не затрагивающего центромеру. — Перицентрическая инверсия — то же самое, но в пределах участка, включающего центромеру. — Инсерция — встройка дополнительного хромосомного материала в какой-либо участок хромосомы. — Делеция — потеря участка хромосомы ХП приводят к изменениям кариотипа (хромосомные дупликации)
3. Генные мутации — изменения генов.
Генные, или точковые, мутации связаны с изменением состава или последовательности нуклеотидов в пределах участка ДНК — гена. Нуклеотид внутри гена может быть заменен на другой или потерян, может быть вставлен лишний нуклеотид и т.д. Генные мутации могут привести к тому, что мутантный ген либо перестанет работать и тогда не образуются соответствующие и-РНК и белок, либо синтезируется белок с измененными свойствами, что приводит к изменению фенотипических признаков особи. Вследствие генных мутаций образуются новые аллели, что имеет большое эволюционное значение.
В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. Если под действием мутации изменяется один нуклеотид, говорят о точковых мутациях. Точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Из-за вырожденности генетического кода могут быть три генетических последствия точковых мутаций: сохранение смысла кодона (синонимическая замена нуклеотида), изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация) или образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA. В соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов.
Б). По проявлению в гетерозиготе:
1. Доминантные мутации.
Доминантные мутации — мутации, проявляющиеся в гетерозиготном состоянии в поколении их возникновения и расщепляющиеся в следующих поколениях.
2. Рецессивные мутации.
Рецессивные мутации — мутации, проявляющиеся, если мутантный ген окажется в гомозиготном состоянии.
B). По уклонению от нормы или так называемого дикого типа:
1. Прямые мутации.
Прямые (первичные) мутации — это мутации, вызывающие отклонение от дикого типа. Обратные мутации — это возвращение к дикому типу.
2. Реверсии. Иногда говорят об обратных мутациях, однако очевидно, что они представляют собой только часть реверсий, поскольку в действительности широко распространены так называемые супрессорные мутации.
Мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной.
Г). В зависимости от причин, вызывающих мутации:
1. Спонтанные, возникающие без видимой причины, т.е. без каких-либо индуцирующих воздействий со стороны экспериментатора.
2. Индуцированные мутации.
Современная точка зрения на причины спонтанных мутаций сформировалась в 60-х годах благодаря выяснению механизмов воспроизведения, репарации и рекомбинации генов и открытию ферментных систем, ответственных за эти процессы. Возникла тенденция объяснять генные мутации как ошибки в работе ферментов матричного синтеза ДНК. Сейчас эта гипотеза общепризнана. Притягательность гипотезы заключается также в том, что она позволяет рассматривать и индуцированный мутационный процесс как результат вмешательства внешних факторов в нормальное воспроизведение носителей генетической информации, т. е. дает единое объяснение причин спонтанных и индуцированных мутаций. Большое влияние на развитие теории мутационного процесса оказало изучение его генетического контроля. Были открыты гены, мутации которых могут повышать или понижать частоту как спонтанных, так и индуцированных мутаций. Эти и другие факты, которые будут рассмотрены далее, — убедительные аргументы в пользу существования общих причин индуцированного и спонтанного мутационного процесса.
Первое объяснение механизма мутационных изменений (генных мутаций и хромосомных аберраций) было предложено в 1935 г. Н. В. Тимофеевым-Ресовским, К. Циммером и М. Дельбрюком на основании анализа радиационного мутагенеза у высших организмов и прежде всего у дрозофилы. Мутация рассматривалась как результат мгновенной перестройки атомов в сложной молекуле гена. Причиной такой перестройки считалось непосредственное попадание в ген кванта или ионизирующей частицы (принцип попадания) или же случайные колебания атомов. Открытие в дальнейшем эффекта последействия ионизирующих излучений показало, что мутации возникают в результате процесса, длящегося во времени, а не непосредственно в момент прохождения кванта энергии или ионизирующей частицы через ген.
Перспективы преодоления этих и других противоречий зарождающейся теории мутационного процесса были намечены в физиологической гипотезе мутационного процесса, высказанной и 1946 г. М. Е. Лобашевым.
Сущность гипотезы М.Е. Лобашева заключалась в том, что «благодаря способности клетки репарировать полученные повреждения становление мутации должно осуществляться в процессе обратимости повреждения, т. е. в процессе восстановления (репарации)». Это означало, что появлению мутации должно предшествовать предмутационное состояние или потенциальное изменение, которое может быть устранено (тождественная репарация) либо реализуется в виде мутации (нетождественная репарация). Для доказательства существования таких предмутационных состояний М.Е. Лобашев, его ученики К.В. Ватти, М.М. Тихомирова и другие в опытах с дрозофилой, облученной рентгеновыми лучами, дополнительно воздействовали на нее высокой температурой, которая сама по себе мутаций практически не вызывала. Мухи, подвергнутые такому комбинированному воздействию, обнаруживали более высокую мутабильность, чем после воздействия только рентгеновыми лучами.
Только эти четыре способа классификации изменений генетического материала носят достаточно строгий характер и имеют универсальное значение. Каждый из подходов в этих способах классификации отражает некоторую существенную сторону возникновения либо проявления мутаций у любых организмов: эукариот, прокариот и их вирусов.
Существуют и более частные подходы к классификации мутаций:
Д). По локализации в клетке:
1. Ядерные.
Ядерные мутации — геномные, хромосомные, точечные.
2. Цитоплазматические. В этом случае обычно подразумевают мутации неядерных генов.
Цитоплазмотические мутации — связанные с мутациями неядерных генов находящихся в митохондриальной ДНК и ДНК пластид — хлоропластов.
Е). По отношению к возможности наследования:
1. Генеративные, происходящие в половых клетках
Если мутации возникают в половых клетках, их называют генеративными мутациями, а если в других клетках организма — соматическими мутациями. Соматические мутации могут передаваться потомству при вегетативном размножении. Генеративные мутации — унаследованные мутации, они возникают в половых клетках, но не влияют на признаки данного организма, а проявляются только в следующем поколении.
2. Соматические, происходящие в соматических клетках
Соматические мутации — мутации в клетках тела. Если изменяются гены в соматических клетках, то мутации проявляются у данного организма и не передаются потомству при половом размножении. Однако при бесполом размножении, если организм развивается из клетки или группы клеток, имеющих мутировавший ген, мутации могут передаваться потомству. Такие мутации называются соматическими.
Очевидно, два последних способа классификации мутаций применимы только к эукариотам, а рассмотрение мутаций с точки зрения их возникновения в соматических или половых клетки имеет отношение только к многоклеточным эукариотам.
Наконец, очень часто мутации классифицируют по их фенотипическому проявлению, т.е. в зависимости от изменяющегося признака. Тогда рассматривают мутации летальные, морфологические, биохимические, поведенческие, устойчивости или чувствительности к повреждающим агентам и т. д. Возможно, это наиболее эклектичный способ классификации, но им довольно часто пользуются в специальной литературе.
В общем виде можно сказать, что мутации — это наследуемые изменения генетического материала. Об их появлении судят по изменениям признаков. В первую очередь это относится к генным мутациям. Хромосомные и геномные мутации выражаются также в изменении характера наследования признаков.
Основные аллели окрасов собак.
Аллели «Агути»
Ряд наиболее распространенных фенотипов окраса определяются серией аллелей, известной как «агути». Этот термин произошел от названия небольшого грызуна с коричневосерой шерстью, которая прекрасно скрывает его от хищников. У псовых соответствующий окрас называют волче-серым (зонарным). Однако, в этом случае, такой окрас позволяет хищнику незаметно подкрадываться к жертве.
В генетике собак термин «агути» позаимствован из генетики грызунов, где ген «агути» хорошо изучен. Выяснено, что исходный (дикий тип) ген «агути» является родоначальником целой серии аллелей. В настоящее время выявлены следующие аллели:
Фенотипическое проявление — Символ
Сплошной черный — As
Доминантный желтый (рыжий) — Ay
Агути (волче-серый) — A
Чепрачный — asa
Черно-подпалый — at
Исходный ген А ( «агути») отвечает за волче-серый окрас диких псовых и большинства, если не всех пород собак, имеющих волчеобразный серый окрас. К таким породам можно отнести серую сибирскую хаски, серого норвежского элькхунда и других собак крайнего Севера. Возможно, что серая немецкая овчарка имеет генотип АА.
В результате мутагенеза ген А дал четыре мутантных аллеля: два доминантных (As и Ay) и два рецессивных (asa и at) по отношению к исходному гену А.
Ген As отвечает за сплошной черный окрас, характерный для многих пород. Этот окрас варьирует от угольно-черного до буро-чёрного. Последний образуется в результате различного числа остаточных агути-подобных волос. Нечистокровные черные собаки часто имеют бурый оттенок. При ближайшем рассмотрении их шерсти можно выявить некоторое количество зонарных волос наряду с чисто черными. Наиболее простое объяснение этому явлению заключается с том, что ген As сам по себе не может индуцировать образование чисто черных волос. Для этого ему требуется наличие достаточного количества модифицирующих полигенов, селекционно отобранных при выведении чистокровных пород. Можно также предположить, что ген As не полностью доминантен, тогда генотип AsAs определяет чисто черный окрас, а генотипы AsAy или AsA — бурый. Агути-подобные волосы могут давать также специфический красноватый оттенок, описанный Литтлем (1957), который предположил, что такой отгонок образуется благодаря неполному доминированию As над Ay и at. Литтл показал, что у особей AsAy красноватый оттенок встречается в основном на боках, голове, шее и конечностях, а особи с генотипом Asat имеют красноватый оттенок только там, где должны располагаться подпалы у особей atat. Такое неполное доминирование вполне возможно. Однако, следует заметить, что статистически это еще не доказано.
Доминантный аллельный ген Ay отвечает за рыжий окрас собаки. Этот ген дает чисто рыжих собак, но при этом обнаруживаются волосы с черными кончиками (т.н. типированные) на голове, плечах, вдоль спины, включая хвост. При наличии значительного количества таких волос можно говорить о соболином окрасе. Поэтому, этот аллель можно назвать также аллелью соболиного или соболе-рыжего окраса. По этой терминологии полностью, рыжие формы можно обозначить как золотисто-соболиные или светло-соболиные. Количество зачерненных волос или степень проявления соболиного окраса варьирует, что обусловлено полигенами, которые наследуются независимо от А. В совокупности, эти полигены известны под названием «затеняющие».
Литтл (1957) предположил, что соболиный окрас проявляется при гетерозиготном генотипе Ayat. Тогда, при скрещивании обоих особей соболиного окраса, потомство должно быть рыжим, соболиным и черно-подпалым в соотношении 1:2:1 соответственно. Экспериментально, примерно так и получалось. Однако, и от чисто рыжих собак также получались периодически черно-подпалые щенки, что говорит о гетерозиготности обоих родителей (Ayat). Это обстоятельство в значительной степени опровергает идею неполного доминирования Ay над at. Можно предположить, что полигены затенения сами по себе так взаимодействуют с генами at и Ay, что в случае гетерозиготы Ayat получается темно-соболиной окрас, а в случае гомозиготы AyAy — рыжий окрас со слабым или незначительным затенением. Но это весьма сомнительно.
Ген чепрачности asa определяет V-образный характер пигментации шерсти по обеим сторонам туловища, как, например, у эрдельтерьера или бигля. На первый взгляд, фенотипы asa и at сходны, но чепрачные особи имеют больше рыжего, чем черно-подпалые, особенно на морде, плечах, боках и конечностях. Однако, большое распространение черного в чепрачном окрасе может напоминать черно-подпалый. Из двух этих окрасов чепрачный наиболее вариабелен.
В типичном случае черно-подпалого окраса (доберманы, например) черный распространяется на всю верхнюю часть туловища, а рыжий ограничен внутренней поверхностью конечностей, грудными отметинами и нижней поверхностью морды. Два характерных пятна расположены над глазами. Для чепрачного окраса характерны возрастные изменения. Так, чепрачные щенки рождаются очень похожими на черноподпалых, с возрастом они светлеют, рыжие отметины увеличиваются по площади, пока молодые животные не становятся истинно чепрачными. Нельзя не отметить интригующий факт, что соболиные собаки подвержены сходным возрастным изменениям. Это явление настолько всеобщее, что и детеныши волка имеют более темный окрас, чем взрослые животные.
Интересно, что у большинства чистокровных пород собак, за исключением эскимосских и скандинавских, аллель дикого типа А отсутствует. Такое отсутствие столь характерно, что кажется преднамеренным. Возможно, что древние селекционеры стремились закреплять такие цветовые вариации, которые, во-первых, отличали бы домашних собак от диких форм и, во-вторых, служили бы символом одомашнивания собак.
Наиболее очевидно, что самым близким аллелем к гену А является аллель asa. В основном А и asa проявляются сходно. Так, у нечистокровных животных наблюдаются все переходы от зонарного (волче-серого) до чепрачного окрасов. Более того, Фокс (1978) предположил, что зонарный и чепрачный окрасы обусловлены действием одного и того же гена. Он скрещивал койота и бигля и в последующих двух поколениях получил в общей сложности 16 животных, которые летом были темно-соболиного окраса, а зимой светлели до светло-соболиного. Однако, по рисункам, представленным Фоксом, потомство было скорее зонарным или светло-чепрачным. В любом случае, это наблюдение показательно, но не убедительно.
Литтл (1957) предположил, что чепрачный и черно-подпалый окрасы обусловлены действием одного аллеля, а различия между ними являются результатом работы модифицирующих полигенов. Однако, в 1976 году Виллис убедительно продемонстрировал различия между генами asa и at и доказал доминирование asa над at. Не очень правдоподобна также и идея о существовании более одного аллеля, кодирующего чепрачный окрас. В этом случае наблюдались бы явные скачкообразные различия, тогда как встречаются постепенные переходы от самых затемненных до самых светлых чепрачных окрасов.
Кажется возможным, что у немецкой овчарки черный окрас кодируется не геном As. Так, Виллис описал черную форму, рецессивную по отношению ко всем остальным окрасам локуса агути. Однако до сих пор неясно, принадлежит ли ген рецессивного черного окраса к этому же локусу или нет.
Ильин (1941) скрещивал волка с черной собакой и получил 7 серых и 6 черных отпрысков. При скрещивании серых потомков между собой было получено 17 серых и 3 черноподпалых, а при скрещивании черных — 12 черных и 3 черноподпалых. Чтобы объяснить полученные данные Ильин предположил существование аллеля черного окраса al, который рецессивен но отношению к А, но доминантен к at. Тогда волк, в данном случае, должен иметь генотип Aal, а черная собака alat. В таком случае, сразу же две случайно отобранные особи имеют очень редкий аллель, что представляется весьма маловероятным. Однако, эти же результаты можно объяснить, если допустить, что волк имел генотип Aat, а собака — Asat.
Аллели протяженности окраса
Предназначение этой серии аллелей может показаться странным, однако оно базируется на том, что эти аллели, как и А — аллели связаны с распределением черной и рыжей пигментации. Возможно, что эта серия аллелей контролирует наличие, отсутствие и протяженность только черной пигментации по поверхности туловища.
К настоящему времени известно три аллеля этого локуса.
Фенотип — Символ
Тигровость — Ebr
Нормальное распространение черного пигмента, обусловленное действием других локусов — E
Полное отсутствие черной пигментации шерсти — e
Исходный ген дикого типа Е отвечает за распространение или продуцирование черного пигмента по поверхности всего тела, например, у чисто черных, чепрачных или черноподпалых собак. Гены Е и е являются мутантными аллелями гена Е. Ген Ebr отвечает за чередование черных и рыжих полос, что характерно для многих пород. Такой окрас называется тигровым. Ген Ebr доминантен по отношению к гену Е. Ген е отвечает за образование только желтого пигмента по всей поверхности тела и препятствует синтезу черного пигмента в волосе, не влияя на черную пигментацию мочки носа, кожи, губ, пасти и век.
Литтл (1957) опубликовал убедительные доказательства того, что ген тигровости (Ebr) доминирует над геном Е, а не наоборот, как считалось раньше. Поэтому, символ ebr для гена тигровости можно считать устаревшим.
Взаимодействие этих трех аллелей с точки зрения доминантности пока окончательно не выяснено. Условно можно допустить, что каждый аллель полностью доминантен по отношению к нижеследующему в представленной таблице (см. выше). Многочисленные вариации экспрессии гена тигровости позволяют предположить о неполном доминировании гена Ebr. Тигровость может варьировать от почти черного окраса, с почти незаметными рыжими пестринами, до песочного, слегка украшенного черными штрихами. С одной стороны, большинство этих вариаций обязано действию модифицирующих полигенов, с другой стороны, усредненный вариант соотношения светлых и темных полос показан для гомозигот EbrEbr. Гетерозиготы EbrE и Ebre имеют отклонения от среднего распределения. Особенно генотип Ebre вызывает уменьшение количества темных полос.
Несмотря на интенсивность пигментации все желтовато-коричневые, желтые, рыжие, красные животные содержат в волосах только желтый пигмент. Реальная степень выраженности окраски зависит от действия модифицирующих генов независимо от того, какой ген присутствует в геноме: Ау или е, или оба вместе.
Литтл (1957) предположил, что наличие маски является доминантным признаком и определяется аллелем из серии Е. Он описал маску как «суперразвитие черного пигмента» обозначив соответствующий ген как Em. Однако, в литературе нет никаких очевидных подтверждений его предположения. В настоящее время благоразумнее относиться с опаской к такому утверждению. Наследуемость маски мы продолжим обсуждать ниже.
Взаимодействие локусов А и Е
Обе серии контролируют распределение черного и желтого пигментов по шерсти у большинства пород собак. Во взаимодействии эти локусы образуют множество хорошо известных окрасов.
Экспрессия аллелей агути зависит от присутствия гена Е, определяющего насколько полно распространяется черный пигмент по шерстному покрову животного под действием какого-либо из генов агути. Такие генотипы могут быть описаны, как:
Окрас — Генотип
Сплошной черный — As-Е-
Ay-рыжий — Ay-Е-
Чепрачный — asa-Е-
Подпалый — atat-Е-
При мутации гена Е к е, весь пигмент волос становится желтым, т.е. прекращается синтез черного пигмента. При этом невозможно становится «выявить экспрессию агутиаллелей, т к. их экспрессия зависит от присутствия черного пигмента. Следовательно, все вышеперечисленные генотипы, при замене E на е, будут давать только рыжий окрас различной степени интенсивности.
Окрас — Генотип
e-рыжий — As-ee
— Ay-ee
— Asa-ee
— atatee
Описывая ранее каждый из сравниваемых локусов, мы выяснили, что у собак существует две независимые системы, отвечающие за рыжий окрас. Обе эти системы ведут себя рецессивно по отношению к черному окрасу. Так, Ау-рыжий окрас получается при скрещивании двух черных особей с генотипами AsAyEE; а е-рыжий — при скрещивании собак любого окраса из агути-серии, но гетерозиготных по локусу Е (Ее). Со сто процентной уверенностью, Ay-рыжий можно отличить от е-рыжего, когда от двух рыжих родителей получаются еще и чепрачный или черно-подпалый окрасы (asa-ЕЕ или atatEE соотв.), как это часто бывает у такс.
У многих особей, окрашенных по типу Ау, обнаруживается варьирующее количество черного пигмента в шерсти, обычно в виде темно-типированных волос на голове, вдоль спины, на плечах и боках. Если такие волосы многочисленны, то получается соболиный окрас. Для Ау-рыжего окраса характерно также наличие черной маски, захватывающей морду и уши. Напротив, e-рыжий окрас не имеет заметных типированных волос или маски. Однако, некоторые Ау-рыжие особи бывают настолько лишены черных волос, что фенотипически неотличимы от e-рыжего окраса.
Если в одной породе встречаются случаи, когда от двух рыжих особей получаются черные щенки, можно утверждать, что генофонд данной породы имеет как Ау, так и е. Например, при скрещивании рыжей собаки с генотипом AyAyEE и рыжей собаки с генотипом AsAsee потомство будет черным (генотип AsAyEe), так как As и E доминантны над Ay и е соответственно. Ау-рыжий не может иметь ген As, следовательно, этот аллель привнесен от е-рыжей особи. Т.о. это доказывает, что скрещиваемые рыжие собаки имеют разное генетическое происхождение своего окраса.
Было бы интересно узнать соотношение описанных двух типов рыжего окраса в одной породе. Однако, на данный момент времени нет никаких систематических данных. Тем не менее, можно высказать некоторые предположения. Скорее всего большинство рыжих особей несут ген Ау. Фактически, любой рыжий окрас, который имеет некоторое количество черных волос на ушах или вдоль спины, скорее образован за счет действия гена Ау, чем е.
Более четко выявить ген е можно: 1) при скрещивании чепрачного или черно-подпалого окрасов друг с другом в разных сочетаниях, если их генотипы asa-Ее или atatEe. Тогда, генотип всех рыжих потомков первого поколения будет ее; 2) при скрещивании рыжих с чепрачными или черно-подпалыми: если получаются черные щенки, то рыжий производитель должен быть As-ее; 3) при скрещивании двух рыжих собак. Как было описано, выше, если получаются черные щенки (AsAyEe), то один из родителей должен быть ее. В данном случае проблема состоит в том, чтобы определить, какой из производителей несет ее.
Литтл (1957) уже обсуждал вопрос о том, как распределяются генетически различные рыжие окрасы по породам. Он предположил, что генотип ее несут следующие породы:
Бигль
Далматин
Английский сеттер
Золотой ретривер
Сеттер гордон
Ирландский сеттер
Лабрадор ретривер
Пойнтер
Пудель
Бернс и Фразер (1966) полагают, что золотистый кокер-спаниель тоже может нести ее, но Литтл (1957) считает, что в этой породе одновременно существуют и Ау-рыжие и ее-рыжие особи. Присутствие обоих генотипов в одной породе может вызвать путаницу у собаководов с начальными генетическими знаниями, которые не представляют, что могут одновременно существовать два генетически различных рыжих окраса. К таким породам Литтл (1957) отнес следующие:
чау-чау
английский спаниель
филд-спанпель
Аллель тигровости, Ebr проявляет свои характерные черты только на рыжих областях, расположение и площадь которых определяется генами серии агути. Ген Ebr индуцирует синтез черного пигмента в волосах, расположенных на рыжих ареалах в виде пигментированных полос или штрихов на рыжем фоне.
Окрас — Генотип
Сплошной черный — As-Ebr—
Тигровый — Ay-Ebr—
Темно-тигровый — asa-Ebr—
Черно-тигровый — atatEbr—
Ген As эпистатичен по отношению к Ebr. Это не удивительно, т.к. As-индивидуумы полностью черные и не имеют рыжих участков, на которых могли бы развиваться тигровины. Однако, в комбинации с геном Ау, Abr экспрессируется.
Несомненно, что у большинства пород тигровый окрас определяется генотипом Ay-Ebr-.
Генотип asa-Ebr-, определяющий чепрачно-тигровый окрас, дает тигровость на рыжих участках типично чепрачного окраса. Расположение полос будет зависеть от типа чепрачности. Если развитие чепрака незначительно, то полосы проявятся на большей части тела, хотя сами по себе они будут темными. Если чепрак развит значительно и охватывает большую часть туловища, то полосы останутся только на ногах и животе, и такая собака будет выглядеть, как очень темно-тигровая.
Генотип atatEbr-, определяющий темно-тигровый окрас, дает полосы, расположенные на подпалах. Фенотип таких животных — наиболее четкое доказательство того, что Ebr работает только на рыжем фоне. Эти собаки, несомненно, черно-подпалые, несмотря на тигровость подпалов, интенсивность которой может варьировать.
Коричневый окрас
Организм млекопитающих продуцирует всего два пигмента — это желтый и черный пигменты. Они содержатся в гранулах, расположенных внутри волоса. Цвет, количество и форма этих гранул и определяют окрас волоса. Черный волос имеет овальные, интенсивно окрашенные гранулы, содержащие черный пигмент меланин. Палевые, рыжие, красные или желтые волосы имеют более мелкие, округлые гранулы, содержащие желтый пигмент.
У животных коричневого окраса (иногда его называют шоколадный или печеночный) или коричнево-чепрачных, или коричнево-подпалых, гранулы в коричневых волосах имеют меньше черного пигмента, чем в черных волосах, и оптически выглядят как коричневые. У тех же животных рыжие волосы не изменены. Очевидно, что мутантный ген, продуцирующий коричневый цвет, действует только на гранулы, содержащие черный пигмент, осветляя их. Для человеческого глаза это выглядит как коричневый. Т.о, здесь мы имеем дело с двумя альтернативными генами, определяющими черный окрас волос (В) и коричневый окрас волос (b). Исходным является ген B. Он доминантен по отношению к мутантному гену b. Ген b изменяет все черные волосы на коричневые, а так же осветляет все пигментированные участки кожи губ, пасти, когтей, мочки носа и радужину глаз.
Сплошной коричневый окрас получается при комбинации гена b с геном As из агути-серки (As-bb). В следующем разделе, касающемся гена ослабления пигментации, вариации коричневого окраса будут обсуждаться более полно.
Т.к. ген b не влияет на пигментацию рыжих районов, то можно всех рыжих собак разделить на два типа: это «черные» рыжие собаки, несущие ген В в генотипе Ау-В- или B-ее, и «коричневые» рыжие собаки, несущие b: Ay-bb или bbee. Эти окрасы внешне не идентичны, так как во втором случае происходит ослабление пигментации кожи и глаз и такие собаки при рыжем окрасе будут иметь коричневую мочку нос, обводку глаз, губ, осветленные глаза и пр.
Вообще, для собак не характерны случаи, когда от коричневых родителей получали черных щенков. Однако, Франклинг (1971) описала случай появления пяти полностью черных пометов при скрещивании семи пар различных коричневых далматинов. При этом она подчеркнула, что владельцы заслуживают полного доверия, а скрещивания происходили под контролем специалистов.
Если это действительно так, то можно предположить, что шоколадный окрас получается не только в результате мутации B в b, но и в результате какой-то неизученной пока мутации другого независимого локуса. Такая ситуация чрезвычайно редка в генетике млекопитающих и известна только у американских норок. Поэтому, приведенные Франклинг сведения не получили всеобщего признания.
Ослабление окраса
Пигментные гранулы в нормально окрашенном волосе расположены достаточно непрерывно по мере роста волос. В основании цвет менее интенсивен. Например, черные волосы имеют интенсивно черный окрас на концах, бледнея к основанию и становясь совсем голубоватыми у корня, благодаря постепенному уменьшению количества гранул по направлению к коже. То же самое происходит и с пигментацией рыжих волос. Они интенсивно пигментированы на концах, бледнея до палевого у основания. При внешнем осмотре более бледные основания полос не видны, но заметны у стриженых или длинношерстных особей.
«Голубизна» голубых собак тоже является результатом уменьшения количества пигментных гранул, однако это обусловлено другими механизмами. Гранулы в волосах таких собак откладываются не непрерывно, а островками или глыбками. Вследствие этого, одни участки волос содержат больше пигмента, другие меньше. В результате этого, для человеческого глаза шерсть, состоящая из таких волос, будет выглядеть серой в случае черного пигмента или палевой в случае желтого пигмента. Различия в диспозиции пигментных гранул обязаны паре генов: D и d. Последний вызывает прерывистое распределение пигментных гранул в волосе. Таким образом, ген D отвечает за интенсивное прокрашивание, а ген d — за ослабление пигментации.
У млекопитающих существует четыре основных окраса, именуемые как черный, голубой, коричневый и лиловый У собак встречаются все четыре окраса и получаются они в результате комбинаций генов b и d с As и Е.
Окрас — Генотип
Черный — As-B-D-E-
Голубой — As-B-ddE-
Коричневый — As-bbD-E-
Лиловый — As-bbddE-
У многих пород собак встречаются и другие окрасы. Если вместо As— в геноме присутствуют atat те же самые четыре основных окраса дают различных двуцветных или подпалых собак.
Окрас — Генотип
Черно-подпалый — atatB-D-E-
Голубо-подпалый — atatB-ddE-
Коричнево-подпалый — atatbbD-E-
Лилово-подпалый — atatbbddE-
Ген d влияет как на черную (коричневую), так и на желтую пигментацию. Следовательно, голубо-подпалый и лилово-подпалый корректнее было бы обозначить как голубо-кремовый и лилово-кремовый. Однако, приставку -подпалый следует сохранять в тех случаях, когда нужно подчеркнуть определенный характер распределения цветов.
Ген d присутствует у огромного количества пород собак и распространенность его зависит от степени популярности голубого окраса Ген b не так распространен, но встречается особенно часто у спаниелей. Действительно, некоторые породы спаниелей всегда гомозиготны по гену о (bb). Это американский и ирландский водяной спаниели и Суссекс. У других пород разрешены как ген В, так и b. Любопытно, что породы, имеющие ген b, как правило, не имеют гена d, следовательно лиловая комбинация (bbdd) крайне редко встречается. Однако, этот генотип формирует основу уникального окраса веймаранской гончей, генотип окраса которой: As-bbddE-.
Большинство любителей-собаководов имеют свои предпочтительные термины для обозначения различных окрасов. Так, коричневый окрас часто называют печеночным или шоколадным. Лиловый окрас настолько редок и необычен, что для него нет общего термина. Окрас веймаранской гончей описывается как серебристый, что является по общему признанию наиболее подходящим термином. Однако, для собаководов полезно знать, что в генетике млекопитающих термин «лиловый» обозначает окрас, обусловленный генотипом bbdd.
В комбинации с генами Ау и Ebr гены В и D и их аллели дают четыре типа тигровых собак.
Окрас — Генотип
Черно-тигровый — Ay-B-D-Ebr—
Голубо-тигровый — Ay-B-ddEbr—
Коричнево-тигровый — Ay-bbD-Ebr—
Лилово-тигровый — Ay-bbddEbr—
Сказанное применимо ко всем формам тигровости, описанным ранее. Сомнительно, чтобы некоторые коричневые и лиловые формы отчетливо распознавались собаководами. В любом случае они были бы необычны.
Ген b действует только на черный пигмент, следовательно, все рыжие фенотипы выглядят обычно, но присутствие этого гена выявляется по изменениям окраски радужины глаз, кожи, когтей. Ген d ослабляет рыжий до кремового, при этом изменения пигментации кожи, глаз и когтей незначительны.
Соболиный окрас имеет генотип Ay-B-D-E-. Интенсивность затенения зависит от наличия модифицирующих полигенов. Причем, черное затенение может быть ослаблено до коричневого под воздействием гена b, тогда получится шоколадно-соболиный окрас. Однако, сомнительно, чтобы такой окрас можно было легко распознать. Многие собаководы определяют такой окрас как желтый или рыжий, особенно, если затенение незначительно и тогда оно не выделяется на рыжем фоне. Соответственно, голубо-соболиный или лилово-соболиный сразу и не отличить от простого кремового или палевого.
Аллели белого окраса, альбинизма
Серия аллелей белого окраса или альбинотическая фундаментально контролирует синтез пигмента по всему телу. Можно только удивляться, почему целая серия генов так названа, хотя альбинизм чрезвычайно редок среди собак. Ответ следует искать в общей генетике окрасов млекопитающих. Альбинизм может быть редок у собак, но он обычен у многих млекопитающих. Локус альбинизма более мутабилен, чем другие локусы. Таким образом, появилась серия аллелей с характерными фенотипами. Более того, эти фенотипы сходны у всех видов млекопитающих, поэтому общие знания хорошо применимы и в случаях собак, так как явление альбинизма у них крайне редко.
Общее количество мутантных аллелей локуса альбинизма неизвестно, но, по крайней мере, три из них установлены.
Фенотипическое проявление — Символы гена
Полное проявление окраса — С
Шиншилла — cch
Голубоглазый альбинизм — cb
Полный альбинизм — c
Все нормально окрашенные собаки имеют ген С, который разрешает экспрессию генов синтеза пигментов. Типично, что аллели, следующие ниже в таблице, разрешают экспрессию все меньшего количества пигмента. Последний ген в таблице полностью запрещает синтез любого пигмента.
Аллель шиншилла (cch) характерен тем, что осветляет рыжий до бледно-желтого, и почти или совсем не влияет на развитие черного пигмента. Похоже, что этот ген присутствует у пород, имеющих бледно-желтый окрас, таких, как норвежский элькхунд. Ген cch осветляет как Ау— рыжих; так и ee-рыжих собак. В частности, наиболее ожидаемым генотипом для золотого ретривера следует признать cchcchee. Окрас таких шиншилловых животных может варьировать от теплого кремового до почти белого. Большинство этих вариаций образуются под действием модифицирующих руфус-полигенов. Литтл же предположил, что значительные вариации этого окраса обязаны существованию более чем одного аллеля шиншилла у собак.
Второй аллель шиншиллы, обозначенный Литтлем, как ce, возможно, отвечает за почти белый фенотип с кремовой тенью вдоль спины, на плечах и голове. Если принять существования этого аллеля, то генотип этих окрасов будет Ау-cece или ceceee. Можно также предположить, что белые вариации окраса у некоторых пород имеют эти же генотипы, например, вестхайленд-уайт-терьер.
Ни один из полностью белых окрасов у собак не является настоящим альбиносным, так как при этом собаки имеют темные глаза. Следует заметить, что возможность существования аллеля типа шиншилла cch основывается более на фенотипических наблюдениях, чем на результатах экспериментальных скрещиваний.
Существование нескольких аллелей шиншиллы можно подвергнуть сомнению, предположив, что не шиншилла, а модифицирующие полигены ослабляют рыжий до палевого во многих случаях. В конечном итоге, пока следует признать достоверным существование только одного аллеля гена шиншилла.
Литтл часто упоминал в своей книге, что ген С не полностью доминантен по отношению к cch. Но это противоречит наблюдениям у других видов животных. Однако, если признать, что в случаях, описанных Литтлем, рыжий окрас ослабляется под действием полигенов, а не cch, тогда неполное доминирование С не удивительно.
Персон и Ашер (1929) описали два очень светло-окрашенных фенотипа со светло-голубыми глазами и тускло-красными зрачками. Ряд исследователей считают, что, по крайней мере, один из них несет аллель альбинизма, который был назван как «голубоглазый альбинизм» (cb). О наследуемости cb очень мало известно. Предложено поместить этот ген между cch и с в представленной выше таблице.
Полный альбинизм, выраженный в чисто белой шерсти, розовых зрачках и просвечивающей красной радужине, крайне редок у собак. Витни (1947) и Литтл (1957) описали таких альбиносов у пекинесов.
Ослабление окраса, сопряженное с розовоглазием
Эта форма ослабления, в целом, обычна для млекопитающих, но редка у собак. Возможно, что одна из форм осветленного фенотипа с голубыми глазами, описанная Персоком и Ашером (см. выше) имела этот ген. Для подобного типа ослабления характерны голубоватый или сероватый окрас и красноватые глаза. Его следует отличать от обычного ослабления до голубого (ген d), когда зрачки остаются темными. Несмотря на уменьшение количества пигмента, «розовоглазое ослабление» не связано с локусом альбинизма, а является мутантной формой (р) гена Р, также отвечающего за развитие интенсивной окраски шерсти и глаз.
При скрещивании между некоторыми из обнаруженных Персоном и Ашером «альбиносов» были получены щенки с темно-окрашенными шерстью и глазами, что может произойти только, если у родителей за окрас отвечали разные независимые гены, продуцирующие розоватые глаза и осветленный окрас шерсти. Например, кроссы между CCpp (розовоглазое ослабление) а cbcbPP (голубоглазое ослабление) дают интенсивно окрашенное потомство (CcbPp).
Красноватые глаза не всегда полностью лишены пигмента (в противном случае они были бы розовыми) и радужина у них часто голубоватая.
К сожалению, Персон и Ашер описали всех обнаруженных красноглазых собак как альбиносов, хотя не все они таковыми были. Рассмотрим возможные комбинации взаимодействие локусов А, С и Р.
Гены cb и p взаимодействуют друг с другом в генотипе cbcb pp и продуцируют псевдоальбиносов. При этом, каждый действует так, чтобы удалить пигмент, разрешенный другим. Глаза в этом случае должны быть розовыми, а шерсть почти или совершенно белая. У животных, гомозиготных по b (bb) гены cb и p могут иметь больший эффект на окрас глаз, чем у имеющих В.
Грифельно-серое ослаблениеО гене грифельно-серого окраса известно очень мало. Он наследуется как доминантный и фенотипически сходен с геном ослабления до голубого (d). Этот ген обнаружен лишь однажды у колли (символ Sg).
Ослабление по типу «пуховки»
Это необычное ослабление в значительной степени промежуточно. Щенячья шерсть таких мутантных черных особей сначала серая, потом, к 6-8 месяцам, становится нормальной черной с легким, светловатым оттенком. Ген не оказывает воздействия на желтый пигмент и цвет мочки носа. Ген «пухового ослабления» обнаружен пока только у колли (символ pp) (Лунд и др., 1970). Как действует и как наследуется этот ген неизвестно.
Мраморный окрас (мерль)
Мерль — название, данное для обозначения окраса, при котором наблюдается неровное прокрашивание в виде более темных и более светлых участков одного и того же цвета. Такой окрас встречается у шелти, колли, такс, мраморных догов (арлекинов). Окрас мерль получается в случае гетерозиготного состояния гена М, который доминантен по отношению к исходному т. Светлые участки получаются при смешении нормальных и осветленных волос, причем, это более заметно на черном фоне, чем на рыжем. В гомозиготном состоянии ген мерль дает сплошной белый окрас, частично или полностью голубую радужину глаза, часто редуцированное глазное яблоко, частичную или полную стерильность.
Генотипы наиболее распространенных мерлевых окрасов следующие:
Окрас — Генотип
Голубой мерль — As-Mm
Голубой/биколорный мерль — atatMm
Рыжий/соболиный мерль — Ay-Mm
При «голубом» или пятнистом мерле районы нормальной и голубой окраски резко различаются. В типичном случае — это голубая собака с черными пятнами неправильной формы, разбросанными на голубом фоне. Сходный эффект можно видеть у черно-подпалого мерля, в большей степени на черпаке и в меньшей — на животе. Ген М — больше влияет на черный/коричневый пигмент, нежели на желтый, поэтому, у чисто рыжих собак мерлевая пятнистость не так явно выражена. Замечено, что ген М усиливает белую пятнистость. Из-за того, что ген мерль в гомозиготном состоянии несет ряд нежелательных признаков, не следует скрещивать гетерозигот, а тем более гомозигот между собой. Логичнее получать мраморных собак, скрещивая мраморных и нормально окрашенных особей. В этом случае получатся как мраморные, так и нормальные щенки. Возможно, что существует более, чем один аллель гена мерль. Это следует из различий в фенотипическом проявлении и результатов скрещивания (Schaible и Brumuiugh, 1976). Одним из них можно считать ген Mh, обуславливающий мраморный окрас дога (арлекин). Этот ген продуцирует белые пятна в гетерозиготном состоянии (Mhm)- Гетерозиготы MhM сходны с ММ по фенотипическим проявлениям (белый окрас, частичная глухота, микрофтальмия). Признаки MhMh, возможно, также сходны с ММ или даже более выражены.
CN — ослаблениеЭтот ген ослабляет как черный пигмент, так и желтый: черный до тускло-серого, а желтый до бежевого или почти белого. Соболиный окрас трансформируется в серебристо серый или почти белый в соответствии со степенью затененности. Мочка носа окрашена в светло-рыжевато-коричневый цвет, что заметно отличает действие гена cn от действия других генов ослабления окраса. Волос CN-щенков имеет более тонкое строение и может даже казаться слегка волнистым. Этот эффект исчезает у взрослых. Ген «CN — ослабления» наследуется рецессивно (символ cn) и является полулетальным, вызывая дефицит нейтрофилов крови, что серьезно влияет на способность индивидуума противостоять бактериальной инфекции. Большая часть щенков, несущих cncn погибает в течении нескольких первых месяцев жизни. «CN — ослабление» пока известно только для колли.
Прогрессирующее поседениеЛиттл (1957) предположил, что прогрессирующее поседение, или посерение, которое наблюдается у некоторых пород собак, происходит благодаря действию полудоминантного гена G. Автор не приводит экспериментальных данных по скрещиванию для подтверждения своего предположения, однако, Витни (1952, 1958) опубликовал результаты, которые указывают на то, что прогрессирующее поседение детерминировано одним геном. Сначала он обозначил этот ген как si, потом согласился с термином Литтля (G). Характер наследования гена (рецессивный, полудоминантный или доминантный) до сих пор еще не ясен окончательно.
Термин «поседение» употребляется в генетике млекопитающих в случае, когда надо отметить появление значительного количества белых волос на теле. В этом случае количество темных волос превосходит количество белых и животное кажется темным, испещренным белыми волосками. Если количество белых волос больше, чем темных, то такой окрас называют чалым. Непонятно пока, то ли это две отдельные генетические формы, та ли различные степени экспрессии одного гена.
Поседение также классифицируется, как постоянное и прогрессивное. В первом случае оно возникает на определенной стадии развития и остается неизменным с возрастом. Прогрессивное поседение возникает постепенно на протяжении значительного периода или всей жизни. Поседение по второму типу связано с геном G.
Щенки, имеющие ген G рождаются черными (или интенсивно окрашенными), потом сереют (светлеют). Гетерозиготы Gg могут изменяться до темно-серо-голубого. В некоторых случаях это происходит достаточно быстро, но, в основном, животные седеют на протяжении всей жизни. Сходные изменения происходят с гомозиготами, но более резко и глубоко.
Поседение начинается в возрасте нескольких недель и со временем окрас становится серо-голубым. Причем, степень выраженности варьирует как от породы, так и от индивидуума. Поседение может происходить равномерно по всей поверхности туловища, а может и частично. Так, например, у бедлингтона верх головы и часть плеч могут стать почти белыми.
В комбинациях с другими аллелями Gg дает слабое осветление окраса, тогда как GG- резкую редукцию пигмента.
Осветление окраса за счет генотипа GG означает, что ген G если не полностью, то частично эпистатичен по отношению к другим генам окраса. Приведем ниже генотипы, представляющие наибольший интерес:
Окрас — Генотип
Серебристый — As-B-D-E-GG
Шоколадно-серебристый — As-bbD-E-GG
Голубо-серебристый — As-B-ddE-GG
Кремово-серебристый — As-B-D-eeGG
Светло кремовый, серебристый — As-B-ddeeGG
Животные имеющие вышеперечисленные окрасы являются серебристыми или чалыми, у которых белые или слабопрокрашенные волосы перемежаются с окрашенными. При этом, если на тела много окрашенных волос, то цвет их легко фенотипически идентифицировать, но, если их мало, определить окрас можно лишь по общему впечатлению. Так, все bb можно идентифицировать по наличию коричневой мочки носа и осветленных глаз. Шерсть может быть с легким коричневатым оттенком. Таким же образом ee-серебристые животные могут иметь бледно-желтый или кремовый оттенок. Все фенотипы можно легко распознать до начала поседения. Следует отметить, что As-B-ddE-GG должны иметь голубой окрас даже до начала поседения благодаря наличию гена d, а не G. Экспрессия гена G может быть не так заметна благодаря аффекту ослабления за счет гена d. Любая порода, где какие-то особи рождаются интенсивно окрашенными, а затем постепенно бледнеют до отчетливо более светлого окраса, имеет в своем генофонде ген G. Очевидно, что он имеется у таких пород, как бедлингтон, керриблютерьер, бобтейл, пудель. Литтл (1957) предложил, что ген G может присутствовать у кернтерьера, денди-динмонт-терьера, скайтерьера и йоркширского терьера. Можно также предположить его наличие у австралийского терьера и австралийского шелкового терьера.
Белая пятнистостьНаличие белого узора на туловище характерно для многих пород собак. Более того, у многих из них характер расположения белых отметин является чуть ли не основным признаком. Теоретически, белые участки рассматриваются как белые пятна, не взирая на количество белого. Раньше этот термин употреблялся только, когда количество белого лимитировано в виде пятен на темном фоне. Теперь термин «белая пятнистость» употребляется более широко и включает также случаи, когда белый преобладает или же собака почти вся белая. При этом белое поле можно рассматривать как одно большое белое пятно.
Распределение и распространение белых отметин следует относительно правильной прогрессии. Так, если расположить пятнистых собак по мере убывания прокрашенных участков, то выявляется следующая закономерность (см. рис.) Сначала белые отметины появляются на груди, лапах, морде и кончике хвоста. По мере увеличения площади белый распространяется на всю грудь, живот, конечности. Долее — на плечи и холку, затем бока и верхнюю часть спины. Постепенно, на теле остаются немногочисленные пигментированные пятна, уменьшающиеся в размерах. На этой стадии также наблюдается закономерность в расположении оставшихся окрашенных пятен. Чаще всего они остаются на боках, голове, особенно вокруг ушей и в основании хвоста.
Экспрессия аллелей белой пятнистости показана в десяти степенях. Степень 0 является показателем того, что собака имеет незначительную пятнистость или вовсе ее лишена. Степень 10 — собака полностью белая (на рисунке не отражено).
Список литературы:
Руководство по разведению животных, пер. с нем., т. 2, М., 1963;
Брюбейкер Дж. Л., Сельскохозяйственная генетика, пер. с англ., М., 1966;
Лобашев М. Е., Генетика, 2 изд., Л., 1967.
Н.А. Лемеза Л. В.Камлюк Н. Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
М.Г.Псалмов «Книга собаковода»
Инге-Вечтомов С. Г. Генетика с основами селекции. – М.: Высш шк., 1989.
Гуттман Б., Гриффитс Э., Сукузи Д. и др. Генетика: Перевод с английского. — Фаир-пресс, 2004.
Генетика окрасов собак. Рой Робинсон перевод: Н. Ю. Адо, 1995.
Интернет ресурсы:
Яндекс Словари
www.slovari.yandex.ru
Википедия
www.ru.wikipedia.org
Общая и Теоретическая Биология
www.afonin-59-bio.narod.ru
.
 16.10.2013,
17389
просмотров.
16.10.2013,
17389
просмотров.